| Zurück | Utricularia (Wasserschlauch) |
|
Wolf-Ekkehard Lönnig 24. April 2011 Die Evolution der karnivoren Pflanzen: Was die Selektion nicht leisten kann das Beispiel Utricularia (Wasserschlauch) Diskussion von Einwänden Auszug aus http://www.weloennig.de/Utricularia2010Diskussion.pdf |
Muss jeder vernünftige, naturwissenschaftlich orientierte Mensch an die Evolution (einschließlich der der karnivoren Pflanzen) glauben? Seit Jahrzehnten wird diese Frage fast uneingeschränkt wie folgt beantwortet 1 "When he [Darwin] finished, the fact of evolution could be denied only by an abandonment of reason.Life Nature Library, "Evolution, p. 10. "It is not a matter of personal taste whether or not we believe in evolution. The evidence for evolution is compelling."Evolution, Genetics, and Man, p. 319, Dobzhansky. "Its essential truth is now universally accepted by scientists competent to judge."Nature and Man's Fate, p. v, Hardin. "The establishment of life's family tree by the evolutionary process is now universally recognized by all responsible scientists."A Guide to Earth History, p. 82, Carrington. "No informed mind today denies that man is descended by slow process from the world of the fish and the frog."Life magazine, August 26, 1966, Ardrey. "It has become almost self-evident and requires no further proof to anyone reasonably free of old illusions and prejudices."The Meaning of Evolution, p. 338, Simpson. "There is no rival hypothesis except the outworn and completely refuted one of special creation, now retained only by the ignorant, the dogmatic, and the prejudiced."Outlines of General Zoology, p. 407, Newman.
Die Antwort auf die Frage, ob nicht nur jeder Naturwissenschaftler, sondern überhaupt jeder vernünftige Mensch an die (Makro-)Evolution und damit auch an die der Karnivoren glauben muss, wäre demnach uneingeschränkt zu bejahen. "Autoritätsbeweise" zählen jedoch in der Forschung nicht, sondern nur echte reproduzierbare naturwissenschaftliche Beweise. Oben haben wir im Detail anhand von 162 Einzelpunkten ausgeführt, dass genau diese Beweise für unsere Thematik die Evolution der karnivoren Pflanzen fehlen und dass überdies die heutigen Evolutionstheorien an der synorganisierten Komplexität etwa des Fangmechanismus (der Saugfalle) von Utricularia scheitern. Fast die gesamte evolutionstheoretische Argumentation zu den karnivoren Pflanzen hat sich Punkt für Punkt als (zumeist mit nachweislich falschen Tatsachen und/oder unzureichender Methodik arbeitende) pure Pseudowissenschaft erwiesen (unverifiable speculations und Zirkelschlüsse). Alle gut informierten Forscher und Denker wissen, dass die entscheidenden Schritte in der postulierten Evolution von Utricularia nicht nur völlig ungeklärt sind, sondern massive Probleme für die Synthetische (und andere Evolutions-)Theorie(n) beinhalten. 1 If arguments fail to resist analysis, assent should be withheld, and a wholesale conversion due to unsound argument must be regarded as deplorable. W. R. Thompson. Für die Entstehung der Lebensformen bietet vielmehr die Intelligent-Design-Theorie einen realistischen Ansatz zu einer überzeugenden Antwort in der Biologie. In diesem Beitrag möchte ich mich kurz mit einigen Fragen und Einwänden zur vorliegenden Arbeit auseinandersetzen, die von allgemeinem Interesse sein dürften. Implizit gehört dazu auch eine Analyse der oft fragwürdigen Strategien und Argumentationsmethoden, mit der heutzutage viele Autoren die (Makro-)Evolution zur Tatsache erklären wollen (die Seitenangaben und die Hinweise auf die "vorliegende Arbeit" bzw. "siehe oben" beziehen sich auf http://www.weloennig.de/Utricularia2010.pdf ). Die persönliche Frage vorweg nach meiner (natürlich von meinen Kritikern 3 ebenfalls wiederholt und massiv in Frage gestellten) Motivation, möchte ich mit dem folgenden kleinen Bekenntnis beantworten: Es ist die Liebe zur Wahrheit, zu meinen Mitmenschen und zu einer möglichst vorurteilsfreien, erkenntnisoffenen Naturwissenschaft, einer Naturwissenschaft, die nicht von vornherein und dogmatisch auf den philosophischen Materialismus festgelegt ist (was heute jedoch generell der Fall zu sein scheint). Was mich betrifft, so schrieb Prof. Dr. Helmut Viebrock 4, Goethe-Universität Frankfurt am Main, einmal dem Studenten W-EL: "Ich halte Sie für ungewöhnlich begabt; Ihre Begabung ist in einer tiefer angesiedelten Gerechtigkeit und Integrität begründet, gefährdet ist sie durch starke Ablenkungen Ich wünsche Ihnen die Möglichkeit, in Konzentration Ihre Gaben und die damit gegebenen Aufgaben zu verwirklichen." Ich hoffe und bete, dass die "starken Ablenkungen" durch zweifelhafte Aussagen zum Thema Evolution mir zugleich die Möglichkeit eröffnet haben (und weiterhin eröffnen), eine bedeutende Aufgabe zu verwirklichen: Aufklärung eines interessierten und für rationale Argumente und Tatsachen aufgeschlossenen Publikums zum Fragenkreis Evolution, intelligentes Design und Schöpfung. 1. Bevor wir zu biologischen Detailfragen übergehen, vielleicht ein Wort zur generellen Frage, ob es wirklich lohnt, sich so ausführlich mit den Behauptungen des (wenn auch stark) im Internet vertretenen Chemieingenieurs M. Neukamm (MN) zu beschäftigen, dem es mehr um die Rechtfertigung seines materialistischen Weltbilds als um die Biologie geht 5 : Dazu sei zunächst festgestellt, dass 'even at the risk of dignifying his highly misinformed further answers' 6 seine Grundhaltung und daraus resultierende Antworten für die vieler weiterer Zeitgenossen und sensu lato auch des Zeitgeistes überhaupt steht und erstaunlicherweise selbst für die führender kirchlicher Kreise, die seine atheistische Publikationstätigkeit > sogar noch materiell unterstützen (siehe unten). Zur materialistischen Motivation ist weiter zu sagen, dass diese genauso auf zahlreiche Biologen zutrifft (Dawkins, Kutschera, Matzke, Meyers, Olson und viele andere). Wenn darüber hinaus auch noch qualifizierte Biologen durch die suggestive Rhetorik eines MN systematisch fehlinformiert und irregeführt worden sind 7 und wenn selbst der Biologe Hubert Rehm, von 2000 bis 2010 mit Kai Herfort Verlagsleiter und Mitherausgeber des Laborjournals, "ein Gelehrter", "ein wandelndes Geschichtsbuch, ein Mathematiker, ein Biochemiker", "ein außergewöhnlicher Schreiber", "ein Stilist", der sich durch "investigativen Journalismus" ausgezeichnet hat 8 , völlig unkritisch akzeptiert, dass nicht nur die Saugfalle des Wasserschlauchs 9 , sondern auch der Flagellenapparat 10 , der Aronstab, das Auge 11 etc., nun von MN und Mitautoren ganz im Sinne der Synthetischen Evolutionstheorie überzeugend erklärt sei 12 , und wie oben schon angedeutet MN inzwischen mit weiteren fragwürdigen und falschen Behauptungen zu den Karnivoren 13 aufgewartet hat, ist es vielleicht nicht ganz unpassend, mit einigen ergänzenden Fakten und Argumenten diesen völlig unrealistischen Eindruck noch weiter zu korrigieren. Und wem es um die Auffassungen von Biologen mit höheren akademischen Graden an biologischen Institutionen geht, so besteht in der vorliegenden Arbeit ebenfalls kein Mangel: Wurden oben doch auch ausführlich die Thesen von promovierten Biologen und (meist auch) Professoren wie Barthlott, Porembski, Müller, Fischer, Juniper, Robins, Joel, Schmucker, Linnemann, Slack, Remane, Taylor, Rivadavia, Kutschera, Blondeau, Jobson und vieler weiterer zitiert und diskutiert, von den Biologen Schneckenburger und Sikorski sowie anderen, die die Behauptungen Neukamms unterstützen, einmal ganz abgesehen. 2. Zur Abwehr des von mehreren ausgezeichneten Biologen vorgetragenen Zirkelschlusseinwands behauptet MN, der Genetiker W-EL hätte den Vaterschaftstest nicht verstanden, denn der Ähnlichkeitsbeweis der Evolutionstheorie sei genauso sicher wie dieser. Dieser Einwand wurde jedoch was die Sicherheit evolutionärer Schlussfolgerungen aus anatomischen und vor allem genetischen Ähnlichkeiten anlangt schon auf den Seiten 56 und 125 behandelt und widerlegt. Ich bitte den interessierten Leser dazu die ausführliche Fußnote auf der Seite 125 mit den zahlreichen Daten genau zu checken. (Kernpunkt nach Rokas et al. (2005): "Despite the amount of data and the breadth of taxa analyzed, relationships among most metazoan phyla remained unresolved. Ähnliche Aussage siehe übernächste Seite. Zur weiteren Bestätigung einige Zitate aus dem Beitrag Bäumchen, wechsel Dich! vom Laborjournal-Chefredakteur Ralf Neumann (nicht zu verwechseln mit MN) 3/2010, p. 49. Nach Hinweis, dass man sich bis vor kurzem einig war, "dass die Flusspferde zu den Paarhufern (Artiodactyla) gehören", stellt er fest, dass der Stammbaum auf der Basis einer ganzen Reihe von 18S-rRNA- und DNA-Sequenzähnlichkeiten jedoch heute völlig anders aussieht: "Die haben plötzlich völlig neue Verwandte. Denn nach den neuen molekularen Daten sind einige Paarhufer wohl näher mit den Walen verwandt als mit den übrigen Vertretern der eigenen Gruppe allen voran eben die Flusspferde." Ein schönes Beispiel für die häufigen Widersprüche zwischen morphologischen und molekularen Stammbäumen. Und er fährt fort (die meisten Hervorhebungen hier und in den folgenden Zitaten von mir, kursiv hier jedoch von R. N.):
"Schön, wenn die Vergleiche so vieler verschiedener Sequenzen immer wieder denselben Baum ergeben. Denn das ist eher selten der Fall. Auch hier ein Beispiel: 1997 schlugen Anna Marie Aguinaldo et al. vor, dass die Vorläufer der Gliederfüßler (Arthropoda) die Panarthropoda, zu denen auch Bärtierchen und Stummelfüßer gehören nicht mit den Ringelwürmern zu den sogenannten Articulata zusammengefasst gehören. Aus ihren umfangreichen 18S-rRNA-Sequenzvergleichen schlossen die Autoren vielmehr, dass im Gegensatz zu diesem Articulata-Konzept die Panarthropoda zusammen mit den Cycloneuralia (zu denen auch die Nematoden gehören) einen ganz anderen Überstamm bilden nämlich die Häutungstiere oder Ecdysozoa. Klar, dass jede Menge Forscher diesen Zwist mit neuen Sequenzanalysen beizulegen versuchten. Jedoch, mit teilweise skurrilen Ergebnissen. 2005 erschienen etwa in ein und derselben Ausgabe von Mol. Biol. Evol. zwei Artikel zum Thema der eine stützte mit neuen molekularen Daten das Ecdysozoa-Konzept (Bd. 22, S. 1246), die Sequenzvergleiche des anderen widerlegten es (Bd. 22, S. 1175). Einer der beiden Artikel muss also zum falschen Schluss gekommen sein. Bis heute lässt sich nicht sagen welcher." 14 Auch die systematischen Widersprüche zwischen den Stammbäumen der Cytochrom B- und Cytochrom C-Gene werden erwähnt. Zwischenfrage: Ist der Vaterschaftstest (genauer der Abstammungsnachweis oder DNA parentage test) tatsächlich so unsicher wie die phylogenetischen Ableitungen? 15 Hier einige weitere Beispiele, die Casey Luskin zusammengestellt hat 16 . Er schreibt am 2. 4. 2011 u. a. 17 : "If one takes a time to read the technical literature in evolutionary biology and systematics, one finds that it's extremely common for phylogenetic trees to contradict one another. In particular, molecular trees often conflict with one another, or conflict with trees based upon morphology. One gene gives you one version of the tree of life, and another gene will yield an entirely different version of the tree 18 . All of this shows that genetic similarity is appearing in places not predicted by common ancestry. An excellent discussion of this problem can be found here . Many papers have reported on this problem. Here is a sample of 10 papers discussing sharp conflicts between molecular trees: James H. Degnan and Noah A. Rosenberg, Gene tree discordance, phylogenetic inference and the multispecies coalescent, Trends in Ecology and Evolution, Vol. 24(6) (March, 2009). This paper notes that: "A major challenge for incorporating such large amounts of data into inference of species trees is that conflicting genealogical histories often exist in different genes throughout the genome." Michael S. Y. Lee, Molecular phylogenies become functional , Trends in Ecology and Evolution, Vol. 14(5): 177-178 (May, 1999). This paper observes that "the mitochondrial cytochrome b gene implied...an absurd phylogeny of mammals, regardless of the method of tree construction. Cats and whales fell within primates, grouping with simians (monkeys and apes) and strepsirhines (lemurs, bush-babies and lorises) to the exclusion of tarsiers. Cytochrome b is probably the most commonly sequenced gene in vertebrates, making this surprising result even more disconcerting." Mushegian et al., Large-Scale Taxonomic Profiling of Eukaryotic Model Organisms: A Comparison of Orthologous Proteins Encoded by the Human, Fly, Nematode, and Yeast Genomes , Genome Research, Vol. 8:590-598 (1998). This paper explains that "different proteins generate different phylogenetic tree[s]" when one looks at the phylogenetic trees of major animal groups. Rokas et al., Conflicting phylogenetic signals at the base of the metazoan tree, Evolution and Development, Vol. 5(4):346-359 (2003). This study recounts conflicts in the metazoan tree, stating: "The robust reconstruction of metazoan history has proven to be a difficult task." Rokas et al., Animal Evolution and the Molecular Signature of Radiations Compressed in Time , Science, Vol. 310:1933-1938 (December 23, 2005). This paper acknowledges that that "[t]he phylogenetic relationships among most metazoan phyla remain uncertain." Again, the problem lies in the fact that trees based upon one gene or protein often conflict with trees based upon other genes. Their study employed the many-gene technique, and yet still found that "[a] 50-gene data matrix does not resolve relationships among most metazoan phyla." Antonis Rokas and Sean B. Carroll, Bushes in the Tree of Life , PLoS Biology, Vol 4(11):1899-1904 (November, 2006). This paper offers a striking admission of deficiencies in the tree of life, acknowledging that "a large fraction of single genes produce phylogenies of poor quality," observing that one study "omitted 35% of single genes from their data matrix, because those genes produced phylogenies at odds with conventional wisdom." What about the technique of simply adding more data? They suggest that "certain critical parts of the TOL may be difficult to resolve, regardless of the quantity of conventional data available." This means that the excuse that problems exist because of "insufficient amounts of available sequence data" is not panning out and more data is not fixing the discrepancies. The paper suggests that "[t]he recurring discovery of persistently unresolved clades (bushes) should force a re-evaluation of several widely held assumptions of molecular systematics." Rokas and Carroll are Neo-Darwinists, and thus one assumption they unfortunately do not re-evaluate is common descent. They suggest the problems can be fixed by using less studied types of molecular characteristics--in short, they appeal to new untried techniques. Perhaps the inability to construct robust phylogenetic trees using molecular data is because common descent is not the answer. Nardi et al., Hexapod Origins: Monophyletic or Paraphyletic? , Science, Vol. 299:1887-1889 (March 21, 2003) This paper finds that the molecular data indicated that six-legged arthropods, or hexapods -- i.e. insects -- are not monophyletic, a striking conclusion that differed from virtually all previous wisdom. As the article stated "Although this tree shows many interesting outcomes, it also contains some evidently untenable relationships, which nevertheless have strong statistical support." Cao et al., Conflict Among Individual Mitochondrial Proteins in Resolving the Phylogeny of Eutherian Orders , Journal of Molecular Evolution, Vol. 47:307-322 (1998) This paper finds that molecular-based phylogenies conflicted sharply with previously established phylogenies of major mammal groups, such as ferungulates, rodents, and primates. The article concludes this anomalous tree "is not due to a stochastic error, but is due to convergent or parallel evolution. Mindell et al., Multiple independent origins of mitochondrial gene order in birds, Proceedings of the National Academy of Sciences USA, Vol. 95: 10693-10697 (Sept. 1998). This paper describes the difficulties encountered when evolutionary biologists have tried to construct a phylogenetic tree for the major groups of birds using mitochondrial DNA. Trees based upon such mtDNA molecules have conflicted with traditional notions of bird relationships. Strikingly, they even find "convergent" similarity between some bird mtDNA and the mtDNA of distant species such as snakes and lizards. The article suggests bird mtDNA underwent "multiple independent originations," with their study making a "finding of multiple independent origins for a particular mtDNA gene order among diverse birds." Finally, a 2009 article in New Scientist titled, Why Darwin was wrong about the tree of life , states: "For a long time the holy grail was to build a tree of life," says Eric Bapteste, an evolutionary biologist at the Pierre and Marie Curie University in Paris, France. A few years ago it looked as though the grail was within reach. But today the project lies in tatters, torn to pieces by an onslaught of negative evidence. Many biologists now argue that the tree concept is obsolete and needs to be discarded. "We have no evidence at all that the tree of life is a reality," says Bapteste. According to the article, the basic problem is that one DNA sequence would yield one tree, while another sequence would yield a different tree: The problems began in the early 1990s when it became possible to sequence actual bacterial and archaeal genes rather than just RNA. Everybody expected these DNA sequences to confirm the RNA tree, and sometimes they did but, crucially, sometimes they did not. RNA, for example, might suggest that species A was more closely related to species B than species C, but a tree made from DNA would suggest the reverse. For the record, this is not the kind of data expected under common ancestry. The article discusses proposals attempting to save common ancestry, largely entailing ad hoc appeals to a process called lateral gene transfer, where bacteria swap genes, thereby muddying any phylogenetic signal. Yet the article observed that conflicts between trees occurs even among higher branches of the tree of life where such gene swapping is not observed to take place, stating: "More fundamentally, recent research suggests that the evolution of animals and plants isn't exactly tree-like either." Among these higher branches, the article found that "The problem was that different genes told contradictory evolutionary stories." This led one scientist to admit that even among these relationships of higher organisms, "We've just annihilated the tree of life." Likewise, as the first paper cited above from Trends in Ecology and Evolution stated, "conflicting genealogical histories often exist in different genes throughout the genome." This is the sort of data that runs counter to the nested hierarchy predicted by common descent. How does this data interface with intelligent design? ID is not incompatible with common descent, and ID is certainly not incompatible with finding patterns of traits that fit within a nested hierarchy. When designers design various structures using 'variations on a theme,' such structures can often be classified as a nested hierarchy. However, designers also can re-use parts in a way that is not required to fit a nested hierarchy. When we find re-usage of parts in a way that cannot be explained by a phylogenetic tree and common descent, this is the sort of data we might expect under intelligent design, but not common descent. And in fact we find much data that is not predicted by common descent. As this short discussion has shown, there's a lot of data that fits into that category. Some other data that does not fit neatly with common descent include: (1) Phylogeny and biogeography often disagree . (2) Phylogeny and paleontology often disagree . (3) Transitional fossils are often missing (or the "predicted" transitional fossils fall apart on closer inspection ). Soweit Casey Luskins' Zusammenstellung und Anmerkungen zu den Widersprüchen phylogenetischer Bäume, die man übrigens noch ausführlich mit den oft unterschiedlichen Ergebnissen ergänzen könnte, die durch die verschiedenen methods of tree construction (distance methods, parsimony methods, maximum likelyhood, Bayesian approaches) bedingst sind. Dazu würde auch eine kritische Diskussion der Problematik der basic assumptions sowie unterschiedlicher Korrekturmethoden gehören (mehrere wesentliche Punkte zum Thema hat Prof. Jeffrey H. Schwartz, University of Pittsburgh, schon sehr gut herausgearbeitet; siehe seinen Beitrag Molecular Systematics and Evolution (2006): http://www.pitt.edu/~jhs/articles/molecular_systematics.pdf (siehe weitere seiner gründlichen, sachkritischen Beiträge unter http://www.pitt.edu/~jhs/publications.html ). Zurück zum Vaterschaftstest: "In a DNA parentage test, the result (called the 'probability of parentage') is 0% when the alleged parent is not biologically related to the child and the probability of parentage typically greater than 99.9% when the alleged parent is biologically related to the child. 19 Nun stelle sich der Leser einmal vor, die Verfechter der Methode des DNA-Vaterschaftstests (Abstammungsnachweis, paternity test, Überbegriff DNA parentage test) müsste statt der erwarteten 99,9% Sicherheit für konkrete Fälle (oder zum großen Teil auch ganz allgemein) Folgendes eingestehen: "Conflicting genealogical histories often exist in different genes throughout the genome, "the mitochondrial cytochrome b gene implied...an absurd family tree for the child (grouping his possible fathers with cats and whales) regardless of the method of tree construction, different proteins generate different family tree[s], i. e. different fathers", "the robust identification of the child's father has proven to be a difficult task." "[t]he relationship to most possible fathers remain uncertain","more data is not fixing the discrepancies, "the child is not monophyletic (i. e. has not only one but several different fathers), a striking conclusion that differed from virtually all previous wisdom, which nevertheless has strong statistical support", "this paper finds that molecular-based family trees conflicted sharply with previously established male parents, "they find convergent similarity between some of the child's mtDNA and the mtDNA of distant species such as snakes and lizards, "we have no evidence at all that the family tree is a reality," .one DNA sequence would yield one tree, while another sequence would yield a different tree, "more fundamentally, recent research suggests that the family tree of the child isn't exactly tree-like either", "the problem was that different genes told contradictory stories to identify the father." Könnte es vielleicht zutreffen, dass MN den gravierenden Unterschied zwischen der 99,9 prozentigen Sicherheit des DNA-Vaterschaftstests sowie der DNA-Forensik (>99%) im Gegensatz zu der oft völligen Unsicherheit phylogenetischer Rekonstruktionen nicht nachvollziehen kann? Stellen wir aufgrund solcher Daten kurz fest: Wenn (a) der DNA-Vaterschaftstest sowie (b) die Ergebnisse der DNA-Forensik genauso unsicher wären wie die phylogenetischen Bäume, dann würde kein Gericht dieser Welt mehr (a) einen genetischen Vater dingfest machen können (ein Kind könnte dann z. B. gleich mehrere genetische Väter haben) und (b) könnten Kriminelle auch nicht mehr annähernd sicher mit dieser Methode identifiziert werden. Der Vaterschaftstest und die DNA-Forensik 20 liegen also auf einer völlig anderen Erkenntnis- und Nachweisebene als die oft widersprüchlichen Phylogenie-Rekonstruktionen. Der Vaterschaftstest liegt innerhalb der genetischen Artgrenzen, die Rekonstruktion der hypothetischen Stammbäume außerhalb. D. h. wir wissen aus der Erfahrung, dass Stammbäume innerhalb der Arten auf Tatsachen beruhen können (zum Artbegriff vgl. Lönnig 2002 http://www.weloennig.de/Artbegriff.html ). Außerhalb der Artgrenzen fehlt dieser empirische (sowie Zeugen-) Nachweis völlig. Als Beispiel eines sicheren Stammbaums sei hier das Fürstenhauses Sachsen-Gotha-Altenburg zitiert:
Vereinfachter Stammbaum des Fürstenhauses Sachsen-Gotha-Altenburg. Quelle: http://upload.wikimedia.org/wikipedia/de/7/75/Sachsen-Gotha-Altenburg.jpg Die evolutionäre Behauptung hingegen, dass das Fürstenhaus Sachsen-Gotha-Altenburg von ausgestorbenen Spitzmäusen abstammt, ist nicht nur wissenschaftlich nicht beweisbar (von den grundsätzlich fehlenden Zeugenaussagen einmal ganz abgesehen), sondern beruht auf der fragwürdigen Voraussetzung der in der vorliegenden Arbeit ausführlich referierten Zirkelschlussmethode mit all ihren widersprüchlichen Ergebnissen (die Methode kurz zusammengefasst: morphologisch-anatomische Ähnlichkeit außerhalb der Arten beruht auf realgenetischer Verwandtschaft, "bewiesen" mit weiteren solchen und anderen Ähnlichkeiten wie biochemisch-molekularen Daten, allen voran mehr oder weniger ähnlichen DNA-Sequenzen). Ich möchte hier noch einmal betonen, dass sich nicht nur die Stammbäume aufgrund der verschiedenen Methoden untereinander häufig widersprechen, sondern dass oft auch noch ein und dieselbe Methode zu widersprüchlichen Ergebnissen führt (weiteres Beispiel: Ableitung der Wirbeltiere von nahezu allen Gruppen der Wirbellosen; vgl. http://www.weloennig.de/mendel13.htm ). 3. Um die (Makro-)Evolutionstheorie vor ihrer Falsifizierung durch Feststellung ihrer unzureichenden Methodik und ihrer widersprüchlichen Ergebnisse zu retten, wird behauptet, dass sie sich auf "Mechanismen und Grundannahmen" stütze, "die unabhängig von der Evolutionstheorie überprüfbar und wohl bestätigt sind. Das der evolutionären Interpretation zugrunde liegende Schema entspricht der hypothetisch-deduktiven "Methode", nach der in allen Naturwissenschaften verfahren wird" (MN, von ihm fett hervorgehoben). Wenn die Schlussfolgerungen und Ergebnisse der Evolutionstheorie tatsächlich unabhängig überprüfbar wären statt auf einem potenzieren Zirkelschluss zu beruhen wieso kommt man dann mit der Theorie laufend zu widersprüchlichen Ergebnissen? Was sollen diese Mechanismen und Grundannahmen sein? Mechanismen: Sämtliche bekannten (und vielleicht noch unbekannten) Mechanismen der "Vererbung und Variation", "die vielschichtigen Mechanismen der Variation, z. B. Genduplikation und Genmutation, der Vererbung und Selektion" etc. bleiben im Rahmen der Mikroevolution. Ich hatte Behe dazu schon auf der Seite 82 oben wie folgt zitiert eine zusammenfassende Aussage, die von MN ignoriert wird: "Indeed, the work on malaria and AIDS demonstrates that after all possible unintelligent processes in the cell both ones we've discovered so far and ones we haven't are at best of extremely limited benefit, since no such process was able to do much of anything. It's critical to notice that no artificial limitations were placed on the kinds of mutations or processes the microorganisms could undergo in nature. Nothing neither point mutation, deletion, insertion, gene duplication, transposition, genome duplication, self-organization, self-engineering, nor any other process yet undiscovered was of much use." Siehe weiter meine folgenden peer-reviewed papers von 2006 http://www.weloennig.de/ShortVersionofMutationsLawof_2006.pdf , 2007 http://www.weloennig.de/Dollo-1a.pdf , 2010 http://www.globalsciencebooks.info/JournalsSup/images/Sample/FOB_4(SI1)1-21o.pdf . Es gibt keine unabhängige Bestätigung der (Makro-)Evolutionstheorie durch Mechanismen (etwa für die Aussage: das Fürstenhaus Sachsen-Gotha-Altenburg stammt von ausgestorbenen Spitzmäusen ab). Die Behauptung, die Theorie sei durch Mechanismen unabhängig überprüfbar und "wohl bestätigt", beruht auf materialistischem Wunschdenken und hat mit den Realitäten nichts zu tun. Zur Selektionstheorie siehe die Ausführungen in der vorliegenden Arbeit und unter http://www.weloennig.de/NaturalSelection.html . Der Kaiser ist nackt. 4. Hypothetisch-deduktive Methode: Nachdem man mit der historisch primär anvisierten und hier tatsächlich naturwissenschaftlich weitgehend beweiskräftigen induktiven Methode (Beweise ad oculus: Reproduzierbarkeit angenommener Evolutionsprozesse (in ähnlicher Form), mutative Erzeugung neuer Arten und höherer systematischer Kategorien, Bildung neuer Organe und Baupläne; vgl. http://www.weloennig.de/NeoA.html ), mit der man ursprünglich die Abstammunsglehre begründen wollte (das erste genetische Journal hieß Zeitschrift für Induktive Abstammungs- und Vererbungslehre; siehe die Diskussion dazu unter http://www.weloennig.de/Popper.html ), systematisch gescheitert ist, beruft man sich als letzten Rettungsversuch auf die hypothetisch-deduktive Methode unter der Vorgabe des methodologischen Naturalismus, mit der man die Gesamtevolution als einzig mögliche Option schlicht und einfach (und völlig unbewiesen) voraussetzt und mit der man auf die oben genannte eigentlich zu erbringende Beweislast praktisch völlig verzichten kann, ohne jemals die Frage geklärt zu haben, ob die Methode in dieser Form zur Wahrheitsfindung in der Ursprungs- und Evolutionsfrage der Biologie überhaupt anwendbar und beweiskräftig ist. Die Methode (als unantastbares und nicht testbares Dogma) schließt von vorherein Intelligenz, Geist, Weisheit, Design und Designer absolut aus und weist sogar die Frage nach der Wahrheitsfindung ab 21 ("Even if all the data point to an intelligent designer, such a hypothesis is excluded from science because it is not naturalistic vgl. http://www.weloennig.de/Die_Affaere1.pdf ).
Mit diesen unabdingbaren naturalistischen Voraussetzungen kann man folgendermaßen vorgehen, wobei die Intuition noch eine ganz besondere Rolle spielt 22 :
Man untersuche mit dieser Methodik (unter grundsätzlichem Ausschluss von Design) etwa die Entstehung der Gemälde in den Galerien des Louvre. Klar kann man damit endlos tolle Evolutionsgeschichten ohne Design und Designer, ohne Genies, Künstler und Architekten, ohne Ziel, Entwurf, Plan und Geist erfinden. Mit den Realitäten allerdings hätte das nichts mehr zu tun (vgl. weiter die Diskussionen unter http://www.weloennig.de/RSGID1.html , http://www.weloennig.de/RSGID2.html und http://www.weloennig.de/RSGID3.html .) Ralf Neumann hat in einem seiner Beiträge Schöne Biologie betitelt "Intuitive Irrtümer" die Bedeutung der Intuition in den modernen Naturwissenschaften inklusive ihrer hypothetisch-deduktiven Methode wie folgt hervorgehoben 23 : [Zitat nach Einstein:] "Höchste Aufgabe der Physiker ist also das Aufsuchen jener allgemeinsten elementaren Gesetze, aus denen durch reine Deduktion das Weltbild zu gewinnen ist. Zu diesen elementaren Gesetzen führt kein logischer Weg, sondern nur die auf Einfühlung in die Erfahrung sich stützende Intuition." Oder der US-Physiker Richard Feynman, der in seinen "Vorlesungen über Physik" die Intuition als absolut notwendig beschreibt, "um aus den Hinweisen die großen Verallgemeinerungen zu finden; um die wunderbaren [...] Gesetzmäßigkeiten hinter den Dingen zu erraten und danach durch das Experiment zu prüfen, ob wir richtig geraten haben." Entsprechend fasste der US-Mathematiker Raymond Wilder 1967 in Science (Bd. 156, S. 605-10) zusammen: "Die Hauptrolle der Intuition ist es, für eine konzeptionelle Basis zu sorgen, welche die Richtung für die weitere Forschung vorgibt. Sie liefert somit einen "educated guess", der sich im Nachhinein als richtig, aber auch als falsch entpuppen kann." Worauf R. Neumann einige Beispiele für intuitive Irrtümer und ihre Verbesserungen aufführt. Wenn aber in der Intuition eines Materialisten zusammen mit der "reinen Deduktion" (Einstein) realistische Ansätze mit Telos (Ziel) und Design auch in der Evolutionsfrage grundsätzlich ausgeschlossen sind, dann können die Lebensformen auch niemals auf eine intelligente Ursache zurückgeführt werden, selbst dann wenn das nachweislich der Fall ist (vgl. dazu Beispiele von Rammerstorfer 2006 und 2010). Der Hinweis auf die hypothetisch-deduktive Methode, die die Abstammungslehre (Makroevolution) unabhängig überprüfbar und rechtfertigen soll, entpuppt sich bei etwas genauerer Betrachtung als pure materialistische Dogmatik. Zusammenfassend können wir also feststellen, dass weder die behaupteten Mechanismen (wie Mutation und Selektion) noch die hypothetisch-deduktive Methode mit ihren materialistischen Grundannahmen und entsprechender Intuition (unter prinzipiell-dogmatischem Ausschluss von intelligentem Design) die (Makro-)Evolution unabhängig überprüfbar machen und bestätigen. Vielmehr steckt in beidem sowohl in den (völlig zu Unrecht) ad infinitum extrapolierten Mechanismen als auch in der Methode die naturalistische Evolutionstheorie als einzig mögliche und damit alternativlose Erklärung für den Ursprung der Lebensformen schon drin. Man vermeidet "die Falsifizierung um jeden Preis" und hat damit in dieser Frage die empirische Wissenschaft aufgegeben (Popper; vgl. wieder http://www.weloennig.de/Popper.html ). Das heißt heute Naturalismus und hypothetisch-deduktive Methode zum Thema Evolutionstheorie. Das folgende Wort von Prof. Walter Höres, Freiburg, gilt auch für den vermeintlich gesicherten wissenschaftstheoretischen Ansatz der heutigen Evolutionstheorie: "[Der Darwinismus] ist und bleibt die größte Zumutung an den menschlichen Geist, die die Neuzeit, ja vielleicht die ganze abendländische Geschichte jemals erlebt ha[t]. Die Herausforderung, die Wunder der organischen Welt damit zu erklären, dass sie "von selbst" entstanden seien." Lässt man hingegen das Dogma der im Prinzip schon vollständig naturalistisch erklärten Makroevolution fallen, dann zeigen sowohl zahlreiche empirische als auch mathematisch-theoretische Ansätze und Schlussfolgerungen die Grenzen der Extrapolationsmöglichkeiten der Mikroevolution-Mechanismen auf (vgl. z. B. Axe 2004, 20010, Dembski und Marks 2009, Junker 2008, 2009a, 2009b, 2010a, 2010b, Lönnig 1976/2003, 2005, 2006, 2010, Luskin 2011 24 ) und sprechen überdies eine deutliche Sprache für intelligentes Design (die Offenheit der wissenschaftlichen Methoden für Design einschließlich der induktiven sowie der hypothetisch-deduktiven ohne materialistische (Ausschluss-)Dogmatik dabei vorausgesetzt; vgl. wieder die Diskussionen unter http://www.weloennig.de/RSGID1.html ff.). 5. Wenn die Aussagen der Evolutionstheorie unabhängig durch Mechanismen und Grundannahmen überprüfbar wären, dann fragt man sich, warum so viele Biologen und andere Vertreter der Evolutionstheorie schon seit Jahrzehnten immer wieder darauf hinweisen, dass die Frage nach dem "Ob" der Evolution "logisch völlig unabhängig und getrennt von der Frage nach dem "Wie" zu behandeln" ist (MN). Denn das "Wie" das gibt man zu weiß man in aller Regel nicht. Als Student ist mir diese Trennung von "ob" und "wie" schon regelmäßig in den 1960er Jahren begegnet. Man weiß zwar nicht wie, wo, wann und warum sich eine komplexe Struktur "von selbst" entwickelt haben soll, aber dass sie sich allein aufgrund der uns bekannten materiellen (physikochemischen) Gesetzlichkeiten entwickelt habe, das sei absolut sicher. Wäre die hypothetische (Makro-)Evolution direkt beobachtbar und reproduzierbar, d. h. würden solche Prozesse direkt vor unseren Augen ablaufen und wir wüssten nur die Ursachen dafür nicht (oder noch nicht), dann könnte man dieser Trennung ruhig folgen (das "Ob" wäre klar, nur das "Wie" wäre noch weiter zu erforschen) und die Frage nach dem "Ob" der Evolution wäre dann "logisch völlig unabhängig und getrennt von der Frage nach dem "Wie" zu behandeln". Tatsächlich aber kennen wir zur postulierten (Makro-)Evolution weder das "Ob" (es ist niemals beobachtet worden etwa die Transformation von ausgestorbenen Spitzmäusen zu Giraffen, Tigern, Bären, Elefanten, Walen und Menschen) noch das "Wie" (kontinuierlich oder diskontinuierlich; die genauen ökologischen, physikochemischen Ursachen und speziellen Abläufe). Der Aktualität halber erinnere ich dazu an einen Gedankengang von Friedrich Andermann 25 , der diesen Punkt im Rahmen einer weiteren Standartentgegnung zur Immunisierung der Evolutionstheorie gegen jegliche Widerlegung ("aber bedenken Sie doch die Wirkung der Zeit") wie folgt veranschaulicht hat: "Rein logisch spielt die Zeitfrage beim Problem der Artbildung überhaupt keine Rolle. Ob sie rasch oder langsam vor sich geht, ist nebensächlich. Vorerst muß sie als Tatsache überhaupt erwiesen sein. Würde uns die Erfahrung etwa zeigen, dass sich ein Schaf im Laufe von zwei oder drei Generationen in ein Nashorn umwandeln kann, so müßten wir das als Tatsache hinnehmen, nicht weniger begreiflich wie jede andere Metamorphose in der Natur (die wir in keinem Falle wirklich "verstehen"). In der individuellen Entwicklung, von der allein es wirkliche Erfahrung gibt, verlaufen die einzelnen Phasen der Metamorphose mit verschiedener Geschwindigkeit. Die Schmetterlingslarve behält ihre Form lange Zeit hindurch bei, ohne auffällige Veränderungen durchzumachen. Tritt sie aber ins Puppenstadium, so verläuft die weitere Umwandlung zur Imago sehr rasch, gleichsam über Nacht. Wüßten wir nicht, daß der flatternde Schmetterling mit den prächtigen Flügeln einmal ein häßlicher Wurm [es gibt auch schöne Raupen, Anm. von WEL] war, und würde nicht diese Verwandlung vor unseren Augen vor sich gehen, wir glaubten bestimmt nicht daran und würden eine solche Behauptung für ein Märchen halten. Wie töricht ist doch der Mensch, der da glaubt, es müsse in der Natur genau so hergehen, wie er es sich vorstellt. Gerade das Beispiel von der Umwandlung der Larve in die Imago zeigt uns sehr deutlich, daß die Natur keinesfalls den Weg der allmählichen Evolution gehen muß, wie ihn die Entwicklungstheoretiker ihr vorschreiben. Und in gewissem Sinne kann man hier sagen, daß zwei Arten auseinander entstehen, ohne daß sie die geringste Ähnlichkeit miteinander haben. Was zwingt uns also zur Annahme, daß eine Art nur aus einer ihr ähnlichen entstehen muß, und - was wissen wir überhaupt davon, wie Arten in der Natur entstehen?" "Wenn nun wirklich in der Natur die Tendenz bestünde, die niederen Arten in die höheren zu überführen, so hätte sie bis jetzt reichlich Gelegenheit gehabt, es uns zu zeigen. Merkwürdig genug, daß sie es während der ganzen geschichtlichen Beobachtungszeit nicht getan hat. Und so ist es von vornherein eine faule Angelegenheit, wenn man zu "unendlich langen" Zeiträumen und zu einer unkontrollierbaren Vergangenheit Zuflucht nehmen muß. Wir sehen immer wieder denselben Versuch, logische Schwierigkeiten dadurch abzuschwächen, indem man das Problem verschiebt und sich auf die Wirkung der Zeit hinausredet." Wie kommt man dann aber zur Behauptung, dass das "Ob" der Evolution als Tatsache so absolut sicher sei als wäre man doch dabei gewesen und als wären bereits alle Ursachen mit Mutation, Isolation, Selektion etc. unter grundsätzlichem Ausschluss von Design im Prinzip schon bekannt (nur die speziellen Abläufe für die konkreten Fälle kenne man noch nicht)? Historisch verhält es sich doch genau umgekehrt. Erst als man mit der Selektionstheorie glaubte, die naturalistische Antwort auf die Frage nach dem "Wie" gefunden zu haben, hat sich die Evolutionstheorie in Biologenkreisen weitgehend durchgesetzt. Obwohl diese Antwort nun in den letzten 150 Jahren aufgrund widersprechender paläontologischer, morphologisch-anatomischer, biochemischer, genetischer und weiterer Daten in weiten Bereichen der Biologie wieder fragwürdig geworden ist, hält man dogmatisch an der (Makro-)Evolution als "erwiesene Tatsache" fest. Mit anderen Worten: In der Ohnmacht, das Postulat eines innerlich denkbar unwahrscheinlichen rein materialistisch deutbaren (Zufalls-) Prozesses, der niemals beobachtet wurde 26 (und der auch nach der Theorie grundsätzlich unbeobachtbar ist; vgl. Dokumentation unter http://www.weloennig.de/Gesetz_Rekurrente_Variation.html speziell die Zitate von Schindewolf, Dobzhansky, Andermann), dem zahlreiche Daten grundlegend widersprechen und der damit naturwissenschaftlich widerlegt ist, aus weltanschaulich-atheistischer Motivation dennoch unbedingt ins Zentrum der theoretischen Biologie stellen zu können, bedient man sich einer petitio principii, indem man die Theorie zur Tatsache erklärt, die dann als solche natürlich "logisch völlig unabhängig" von den vielen Detailfragen besteht und in deren Sinne man alles weitere "deduziert": "Eine Petitio principii (lat. "Inanspruchnahme des Beweisgrundes"; engl. "begging the question"), auch Circulus in demonstrando, ist ein Scheinbeweis, bei dem eine Behauptung durch Aussagen begründet wird, welche die zu beweisende Behauptung schon als wahr voraussetzen. Dies kann zum einen explizit geschehen, wenn die Behauptung als Konklusion eines Arguments vorliegt, in dem sie selbst als Prämisse vorkommt, zum anderen implizit, indem die Konklusion kein expliziter Bestandteil des Arguments ist, sondern stillschweigend angenommen wird" http://de.wikipedia.org/wiki/Petitio_principii (dort weitere Ausführungen zum Thema). Da die grundlegenden naturwissenschaftlich-experimentellen Beweise für die Allgemeine Abstammungslehre (Makroevolution) niemals geliefert wurden, kann man die oben postulierte logische Trennung von "ob" und "wie" keineswegs einfach als unantastbar und sicher hinnehmen. Wenn das Faktorensystem des Neodarwinismus (=Synthetische Evolutionstheorie) als rein materialistische Erklärung zur Entstehung etwa der Karnivoren sowohl selektionstheoretisch als auch mutationsgenetisch weitgehend scheitert, dann lässt das selbstverständlich die Frage nach der Berechtigung der ohnehin unbewiesenen Makroevolution nicht einfach unberührt (wie es die Protagonisten des Materialismus zur vollständigen Immunisierung ihres Weltbilds gegen jedwede wissenschaftliche Falsifikation gerne hätten). Da historisch die Anerkennung der Evolution als Tatsache in weiten Kreisen der Biologen aufgrund der vermeintlich naturwissenschaftlichen Erklärungs- und Beweiskraft der Selektionstheorie erst nach 1859 (Darwins Origin) folgte, stellt sich angesichts des Versagens der damaligen und heutigen Theorien vor den biologischen Tatsachen (z. B. Kambriumproblem, Entstehung komplex-synorganisierter Strukturen und Organe, Entstehung neuer Information, irreducible and specified complexity etc.) die Frage, inwieweit der Nachweis der Untauglichkeit nun den Umkehrschluss zulässt und damit auch die Allgemeine Abstammungslehre (Makroevolution) wieder in Frage stellt. Eine Veranschaulichung: Wieso sollte man einer nachdrücklich vorgetragenen Behauptung Glauben schenken, dass einst eine elegante Brückenkonstruktion über einen 40 km breiten und 2 km tiefen Abgrund existierte (sagen wir von einem Alpenberggipfel zu einem anderen), wenn alle heutigen Brückenbau-Versuche angesichts einer derart gigantischen architektonischen Aufgabe und Problematik versagen (d. h. obwohl man nicht sagen kann, "wie" man eine derartige Kluft architektonisch und statisch überhaupt überbrücken konnte) und es überdies keinerlei unbestreitbare Unterlagen und stichhaltige Beweise dafür gibt, dass die hypothetische Brücke tatsächlich jemals existierte. Der weltanschaulich bedingte Hinweis, dass an der Frage, "ob" sie jemals existiert habe, keinerlei Zweifel möglich sei und dass das "Ob" "logisch völlig unabhängig und getrennt von der Frage nach dem "Wie" zu behandeln" sei, dürfte in diesem Zusammenhang wenig überzeugend wenn nicht gar absurd sein. Markus Rammerstorfer (2006, 2010) hat übrigens sehr überzeugend ausgeführt, dass die Bringschuld und Beweislast historisch bei den Vertretern der Evolution liegt. Ebenso Junker 2008, p. 77/78 (kursiv von ihm):
"Wer eine Behauptung aufstellt, die gegen alle bisherigen Erfahrungen (auch in der Biologie in den Fragen der Evolutionsmechanismen) steht, trägt die Beweislast. Eine solche Behauptung ist die Entstehung von Design-Kennzeichen ohne Designer, bei Lebewesen also die Behauptung, die Entstehung der Lebewesen könne allein durch physikalisch-chemische Vorgänge erfolgen. Solange es nicht gelingt, diese Behauptungen zu beweisen oder wenigstens plausibel zu machen, ist es ganz und gar vernünftig, mit einem Designer zu rechnen. Evolutionstheoretiker verweisen an dieser Stelle auf unsere noch unzureichenden Kenntnisse und auf anzunehmende zukünftige Lösungen. Beides ist nicht widerlegbar (wenn auch nur Lückenbüßer). Sich auf "zukünftige Forschungsergebnisse" zu berufen führt nicht weiter. Auch der Design-Ansatz und darüber hinaus jede beliebige andere Position könnte sich auf zukünftige Evidenzen berufen." Die Frage nach dem "Wie" kann die Frage nach dem "Ob" also direkt und unmittelbar tangieren, zumal wenn das "Ob" niemals im postulierten Sinne nachgewiesen wurde. Die vorliegende Utricularia-Arbeit wirft die Frage nach einer Alternative zur materialistischen Makroevolutionstheorie auf. Nun könnte man vielleicht einwenden, dass die unterschiedlichen Lebensformen ja nicht schon ewig existiert haben und folglich irgendwie entstanden sein müssen und dass deshalb die Makroevolution ohnehin die einzig mögliche Antwort auf die Ursprungsfrage sei. Dazu ist zu sagen, dass schon mehrere naturalistische und theistische Alternativtheorien zur Makroevolution (im Sinne von Makrotransformationen wie Spitzmäuse über viele Zwischenstufen und Generationen zum Fürstenhaus Sachsen-Gotha-Altenburg) vorgeschlagen wurden. Hier einige Beispiele ohne Anspruch auf Vollständigkeit: Unabhängige Schöpfungsakte (Göppert (1865) 27 , Genealogie der Urzellen (Wigand 1872), Emikationstheorie (Nilsson 1953), Genomic Potential Hypothesis (Schwabe 2001), direkte Erschaffung polyvalenter Grundtypen (Junker und Scherer 1986, 2006, ähnlich Lönnig im Sinne des genetisch-plasmatischen Artbegriffs 1986, 2010). Mehrere weitere Autoren lassen die Frage als noch weitgehend unbeantwortet offen (z. B. Kerkut 1960/1965 mit der Möglichkeit einer nichtevolutionären Entstehung der Lebensformen). Verschiedene Ansätze, die direkt auf die Genesis zurückgehen, sind dabei noch gar nicht berücksichtigt. 6. Meine Darstellung der Karnivoren gehe von einem "antiquierten Verständnis von Evolution" (MN) aus. Da diskutiere ich ausführlich die neuesten und modernsten molekularen Ergebnisse inklusive evolutionärer Methoden könnte es sich hier vielleicht um den Einsatz pejorativer Begriffe handeln, um den Intelligent-Design-Ansatz beim evolutionsgläubigen Publikum von vornherein in Misskredit zu bringen? 7. MN bestreitet, dass er bis auf eine Ausnahme mit "falschen Tatsachen" gearbeitet habe: Sein Einwand zu Utricularia multifida (früher Polypompholyx multifida) in meinen Worten zusammengefasst: Die Beiträge von Francis E. Lloyd (1936, 1942) seien alt und daher wenig glaubwürdig, die von Reifenrath et al. aber sei neu (2006) und bediene sich modernster und qualifizierter elektronenmikroskopischer Untersuchungen und sei daher maßgebend. Nun ist F. Lloyd so etwas wie der Einstein unter den Karnivorenforschern 28 . Er hat seine erste (falsche) Hypothese ("that in Polypompholyx [U. multifida] the door acts as a simple valve and is incapable of contributing to the sustention of a low pressure of water within the trap"), die er nach Studien an totem (in Spiritus eingelegtem) Material aufgestellt hatte, später revidiert: An lebendem Material in Australien fand er bei U. multifida die im Prinzip gleiche hochkomplexe Saugfalle wie bei allen anderen Utricularia-Arten und hat diese Ergebnisse entsprechend genau beschrieben und auch mit Fotografien dokumentiert. Reifenrath et al. aber beachten diese Arbeiten erst gar nicht und stellen (ebenfalls wieder an "totem", d. h. präpariertem Material) nichts weiter auf als die evolutionstheoretisch motivierte, aber völlig unbewiesene Hypothese U. multifida "might not function with a low pressure-suction movment" und sei ein Bindeglied zu Genlisea (vgl. http://www.weloennig.de/Polypompholyx.pdf und in der vorliegenden Arbeit pp. 62-64). Solange noch detaillierte Tatsachenbeschreibungen und -feststellungen in den Naturwissenschaften einen höheren Stellenwert haben als völlig unbewiesene Evolutionshypothesen, gilt selbstverständlich das Wort von Francis Ernest Lloyd (es sei denn man könnte nachweisen, dass seine Beschreibung in den entscheidenden Punkten falsch gewesen wäre und dass sich genauso alle folgenden Forscher, die die Saugfalle von U. multifida selbst untersuchten, ebenfalls geirrt haben siehe die Autoren unten). Wer hingegen eine äußerst fragwürdige Hypothese wie die von Reifenrath et al. als wissenschaftlichen Tatsache behandelt, geht von einer "falschen Tatsache" ("false fact") aus bewusst oder (in den vorliegenden Fällen vermutlich meist) unbewusst. Was nun die Genauigkeit der Forschung and die Glaubwürdigkeit der "Alten" anlangt, so möchte ich doch daran erinnern, dass wir noch nie so viel Betrug und Täuschung in der Wissenschaft feststellen mussten wie in den letzten sagen wir einmal drei Jahrzehnten. Mein Eindruck ist, dass (vor allem von Haeckel und einigen weiteren Evolutionisten einmal abgesehen) die "Alten" bei ihrer Arbeit in der Regel von einem wesentlich höheren Arbeitsethos getragen wurden als manche zeitgenössischen Forscher (vgl. zum Thema Betrug und Täuschung in der Wissenschaft nur einmal die zahlreichen kritischen Beiträge allein in den letzten Ausgaben des Laborjournals bis April/Mai 2011). MN stellt zusammenfassend zu Reifenraths Hypothese fest: "Um REIFENRATH [2006] zu "widerlegen" zitiert LÖNNIG ausschließlich ältere Arbeiten, ja z. T. sogar Arbeiten aus dem Jahr 1942. Das wäre ungefähr so, als wollte man die 1950 erfolgte Strukturaufklärung des Vitamin B12 anhand von Arbeiten aus dem Jahr 1886 widerlegen." (Schriftbild von MN.) Als wären gründliche und saubere (und in diesem Falle nach weiteren präzisen Untersuchungen materialbedingt sogar revidierte) naturwissenschaftliche Tatsachenbeschreibungen aus dem Jahre 1942 automatisch falsch oder unzuverlässig/fragwürdig (von weiteren Autoren in den folgenden Jahren einmal ganz abgesehen) und völlig unbewiesene evolutionistische Hypothesen von 2006 automatisch richtig und zuverlässig! Legen wir die 64 Jahre zwischen den beiden Publikationen einmal in die Zukunft: Wären dann alle noch so gründlichen, besten und genauesten naturwissenschaftlichen Tatsachen-beschreibungen von 2011 die Kontinuität der heutigen Welt vorausgesetzt im Jahre 2075 nichts mehr wert? Und dieses fragwürdige Beurteilungskriterium 29 kann man natürlich endlos weiter extrapolieren für die Jahre 2139, 2203, 2267 etc. (vgl. dazu weiter die Fußnote 4 auf der Seite 4 oben). MNs Vitamin B12-Beispiel ist übrigens aus mehreren Gründen völlig verfehlt: Abgesehen von dem untergeordneten Punkt, dass die Strukturaufklärung des Moleküls nicht 1950, sondern 1955 durch Dorothy Crowfoot Hodgkin erfolgte (nach Vorarbeiten 1954: "In 1955, Dorothy Hodgkin took the first X-ray diffraction photographs of vitamin B-12" http://www.rcuk.ac.uk/ResearchCareers/dhpa/Pages/AboutDorothy.aspx . Reference: Hodgkin DC, Pickworth J, Robertson JH, Trueblood KN, Prosen RJ, White JG. 1955. The crystal structure of the hexacarboxylic acid derived from B12 and the molecular structure of the vitamin. Nature 176:325-328, sind dabei folgende Punkte zu berücksichtigen: Im Jahre 1886 hatte man noch nicht die geringste Ahnung davon, dass das Vitamin B12 überhaupt existiert. Erst kurz vor 1920 entdeckte George H. Whipple, dass man blood loss bei Hunden wie folgt behandeln kann: " he discovered that ingesting large amounts of liver seemed to most-rapidly cure the anaemia of blood loss [in dogs]. "[D]ie Suche nach der essenziellen Komponente dieser Heilmethode [führte] schließlich 1926 zur Beschreibung eines auch beim Menschen wirksamen "Antiperniziosa-Faktors" durch die beiden US-amerikanischen Ärzte George R. Minot und William P. Murphy die dafür zusammen mit Whipple 1934 den Nobelpreis für Medizin erhielten". Soweit mir bekannt, machte auch in den 1920/30ger Jahren noch niemand einen Vorschlag zur Strukturaufklärung vom Vitamin B12, zumal damals das Vitamin noch gar nicht isoliert worden war und man noch nicht genau wusste, was hinter den Behandlungserfolgen eigentlich steckte. "Die Isolierung des eigentlichen Wirkstoffs dagegen, des Vitamins B12 (Cyanocobalamin), gelang in kristalliner Form erst 1948, und das unabhängig voneinander zum einen einem Team US-amerikanischer Biochemiker um Karl A. Folkers , zum anderen einem britischen Forscherteam um den Chemiker E. Lester Smith . 1955 schließlich konnte die britische Biochemikerin Dorothy C. Hodgkin mit Hilfe der Röntgenbeugung an Vitamin-B12-Einkristallen auch deren Molekülstruktur aufklären, wofür sie u. a. 1964 mit dem Nobelpreis für Chemie geehrt wurde. Die darauf aufbauende Totalsynthese des Vitamins B12 gelang 1972 Albert Eschenmoser und Robert B. Woodward, und noch heute gilt Vitamin B12 damit als eines der größten jemals in einem Labor Moleküle" (vgl. totalsynthetisierten http://de.wikipedia.org/wiki/Cobalamine#Geschichte (2011) und http://en.wikipedia.org/wiki/Vitamin_B12#History (2011); siehe auch Römpp Chemielexikon, 9. Auflage 1995, Bd. 6 p. 4946; man könnte hier vielleicht noch ergänzen, dass im Stoffwechselgeschehen des Menschen die Umwandlung von Cyanocobalamin in Methylcobalamin und Adenosylcobalamin als "die eigentlich biologisch wirksamen Coenzym-Formen des Vitamins" erfolgt siehe http://de.wikipedia.org/wiki/Cobalamine .) Mir ist jedoch nicht bekannt, dass die Tatsachenbeschreibungen aus dem Jahr 1926 zu den Leberextrakten falsch gewesen wären oder etwa das Folgende: "In 1928, the chemist Edwin Cohn prepared a liver extract that was 50 to 100 times more potent than the natural liver products. The extract was the first workable treatment for the disease [pernicious anaemia in humans]. Und die Befunde und Tatsachenbeschreibungen von Cohn sind sogar noch etwas älter als die von F. E. Lloyd zum Fangmechanismus von Utricularia multifida. Natürlich wäre es keineswegs verkehrt, zur weiteren Bestätigung des Saugmechanismus von U. multifida (und damit vielleicht der zusätzlichen Widerlegung der sachlich unbegründeten Hypothese von Reifenrath et al.) und vor allem zur Vertiefung der Beobachtungen und Befunde der bisherigen Autoren samt fotografischer Dokumentation von Lloyd noch weitere unabhängige Untersuchungen an lebendem Material von U. multifida mit ihren besonderen Charakteristika vorzunehmen; sehr erfreulich wären dazu auch Zeitrafferaufnahmen. Mein bisheriger Eindruck von U. multifida ist, dass es sich bei dieser Art trotz einiger 'urtümlicher' Merkmale um eine hochspezialisierte Form handelt. Das dürfte jedoch eine ausführliche Behandlung für sich wert sein. Im Übrigen gehen auch die meisten neueren Autoren davon aus, dass die Beschreibung von Lloyd (1936, 1942) völlig korrekt ist (Benzinq 1987: "ultra-sophisticated suction-traps (Utricularia sensu lato)", Juniper et al. 1989, ebenso Taylor 1989/1994, Slack 1986/2000, 2006, J. und P. Pietropaolo (2001) 30 die letzteren vier mit eigenen Studien zur Falle) wie z. T. oben und in http://www.weloennig.de/Polypompholyx.pdf schon zitiert (vgl. auch implizit die zahlreichen neueren Autoren, die auf den Seiten 17-19 der vorliegenden Arbeit aufgeführt wurden und werden), im Kontrast von MNs Behauptung, dass ich ausschließlich auf ältere Arbeiten verweise. Zur Ergänzung siehe z. B. H. Lambers, F. S. Chapin und T. L. Pons 2008, p. 535: " detail of the trap of Utricularia multifida (bladderwort) with bladder-like suction traps"; R. F. Evert with the assistance of S. E. Eichhorn 2006, p. 465: " suction traps (Utricularia, Biovularia, Polypompholyx) " W. X. Schulze 2010: "Polypompholyx is an Australian species with suction traps in the soil 31 ; P. D. Johnson, under the advisement of D. Inouye (2005, pp. 20, 21, and 23) unter "Spring-traps": Polypompholyx 32 . Britannica Online Encyclopaedia (2011): "Polypompholyx (two species; Australia) are very similar to Utricularia and also trap their prey by means of highly specialized bladders 33 . Honda (2011) 34 beschreibt unter den Überschriften "SUCTION TRAP" und "TRAP OPERATION" die bladderworts und deren trap setting, triggering, resetting, glands, water pumping mechanism, digestion und erwähnt dabei auch mehrere offenen Fragen sowie zwischendurch Loyds Kommentar: "But most to be wondered at are the traps which present an astounding degree of mechanical delicacy depending on a fineness of structure scarcely equaled elsewhere in the plant kingdom." In der Literatur führt er u. a. die Arbeit von Reifenrath et al. (2006) auf, erwähnt aber nicht deren fragwürdige Hypothesen (U. multifida "might not function with a low pressure-suction movement" und hätte möglicherweise einen "primordial (non-suction) trapping mechanism similar to that of the eel-traps of the closely related genus Genlisea"). Außerdem wird in keiner (anderen?) der mir bisher bekannten Darstellungen und Detailbeschreibungen des Fangmechanismus von U. multifida die unbegründete Hypothese vermittelt, dass sie eine "permanent geöffnete Tür" (MN) besitzen könnte. Selbst Lang (1901), der wie Darwin 35 und andere den Saugmechanismus von Utricularia noch nicht erkannt hatte und der meinte, dass Tiere in den Fangapparat "kriechen", stellt zu U. multifia (Polypompholyx multifida) Folgendes fest: "Die Blasen sind dadurch ausgezeichnet vor anderen Schlauchblättern, dass sie keine freie Eingangsöffnung besitzen, sondern eine trichterförmige Eingangsöffnung, welche durch eine Klappe verschlossen ist, die auf einem hufeisenförmigen Widerlager ruht."
M. S. Reut and R. Jobson schließen sich den bisher zitierten Autoren (abgesehen von Lang 1901) zum Thema "Bladder-trap function in the subgenus Polypompholyx" 2010, pp. 158/159 und 160 mit folgenden Feststellungen und Argumenten völlig an und weisen die Hypothese von Reifenrath et al. wie folgt zurück (emphasis again added): "All members of the genus Utricularia possess modified leaves that form suction-bladders for prey capture (Lloyd 1942; Juniper et al. 1989), which are all thought to be active in their pumping of internal fluid to the exterior during trap resetting (Lloyd 1942; Juniper et al. 1989). These metabolically expensive bladders have been implicated as key adaptive innovations during radiation of the genus (Jobson et al. 2004; Laakkonen et al. 2006; Albert et al. 2010). However, U. multifida and U. tenella, which are species circumscribed to the section Polypompholyx (Fig. 1) (Taylor 1989), share a distinctly robust bladder-trap form (see above), with trigger hairs that are either highly reduced (Lloyd 1942; Taylor 1989), or not present (Reifenrath et al. 2006). The thick sidewalls and door, along with the funnel-like form of the entranceway, lead Reifenrath et al. (2006) to speculate that there may be a corresponding absence of bladder-trap function, and the possibility of a passive eel-trap function as found in Genlisea, the sister lineage of Utricularia (Reut 1993a; Jobson and Albert 2002). These characters also lead Lloyd (1932) to the same conclusion as that of Reifenrath et al. (2006). However, after closer examination of live material, Lloyd (1942) found these bladders to function normally. Furthermore, the passive bladder-trap hypothesis is also confounded by the presence of nonfunnelled entranceways and highly developed trigger hairs in the closely related species U. westonii (sect. Tridentaria), which is also assigned to the subgenus Polypompholyx sensu Taylor (1989). Within the section Pleiochasia, bladder-traps seem to function normally, with studies involving U. monanthos having reported the typical suction function (Lloyd 1942; Sydenham and Findlay 1972; Fineran and Lee 1974; Fineran and Gilbertson 1980; Juniper et al. 1989). There is also ecological evidence for normal function, with a field study finding strong similarity in the array of entrapped prey organisms between U. uniflora R.Br. (sect. Pleiochasia) and U. uliginosa Vahl (sect. Oligocista A.DC., subg. Bivalvaria), growing in sympatry across two populations (Jobson 1998; Jobson and Morris 2001). Überdies hat Allen Lowrie, (Duncraig near Perth, Western Australia) erst kürzlich eine vielleicht kann man sagen Wasserform von Utricularia multifida (Polypompholyx multifida) entdeckt, genauer gesagt nennt es sie P. aff. 36 multifida die sich durch folgende Merkmale auszeichnet: "Plants growing as affixed aquatics in 30 cm depth of water, scapes buoyant and erect, flotation provided by the scapes' large celled central core, 3-6 pink flowered inflorescence held erect to 10 cm above the water surface. Mir ist nicht bekannt, dass bei irgendeiner der rezenten Wasserformen spätestens nach Lloyds bahnbrechender Arbeit von 1942 auch nur ansatzweise spekuliert wurde, dass sie sich vielleicht doch nicht durch aktive Saugfallen auszeichnen würde 37 . Nebenbei bemerkt entspricht die Entdeckung der Wasserform U. aff. multifida (oder nach Lowrie P. aff. multifida) dem Ansatz und den Erwartungen der These polyvalenter Grundtypen (vgl. pp. 80, 122, 124). 38 Forschungsaufgabe: Genetisch nachzuprüfen ist unter anderem, ob diese neu entdeckte Linie mit den bisher bekannten Formen von Utricularia multifida und vielleicht auch U. tenella fertil ist in diesem Falle würde sie gemäß dem genetischen Artbegriff tatsächlich zur selben polymorphen Art U. multifida gehören. Nach allem was man bisher nur sagen kann, geht MN im Zuge seiner evolutionstheoretischen Argumentation mit der Voraussetzung, dass Utricularia multifida sich durch absence of bladder-trap function etc. auszeichne (vgl. dazu oben pp. 2, 58, 64, 65, 96, 106, 120, 187) von einer falschen Tatsache (false fact) aus. Im Übrigen hat er nicht widersprochen, dass er mein Polypompholyx-Paper http://www.weloennig.de/Polypompholyx.pdf vorher schon kannte. Utricularia multifida aus http://mycorrhizas.info/nmroots/utraps.jpg Siehe auch Lambers et al. 2008, pp. 534/535: " detail of the trap of Utricularia multifida (bladderwort) with bladder-like suction traps.
Oben: Falle von Utricularia multifida nach Slack 2000, p. 180 (Originalzeichnung von ihm nach eigenen mikroskopischen Studien; unter Kapitel 9 The Bladder Traps; deutsche Ausgabe: Die Saugfallen. "Detailed cross section of bladder to show structure. Passend spricht er von der "Polypompholyx multifida bladder trap". Slack weist im Text (p. 180) u. a. auf Folgendes hin: "The trap [of Polypompholyx] differs in some way from those of Utricularia" worauf die recht genaue Beschreibung der Unterschiede und Eigentümlichkeiten folgt. Zu den Unterschieden gehört jedoch nicht das Fehlen des Saugmechanismus. (Teil-)Definition zu bladder trap: "The bladders ( vesicula ) pump ions out of their interiors. Water follows by osmosis , generating a partial vacuum inside the bladder http://en.wikipedia.org/wiki/Carnivorous_plant . Der genaue Beobachter Francis E. Lloyd hat schon 1936, p. 104, dazu das Folgende festgestellt: "When the trap is set, the walls are concave, more especially the side walls, the top being less responsive (siehe weitere Punkte unten und unter http://www.weloennig.de/Polypompholyx.pdf ). Taylor (1989/1994, pp. 79-84) findet gemäß seinen Untersuchungen, Beschreibungen und Zeichnungen definitiv ebenfalls suction traps bei Utricularia multifida und ebenso bei U. tenella (praktisch die Zwergform von U. multifida), wie im eben zitierten Link ausführlich dokumentiert (vgl. dazu auch Originalzeichnung von J. und P. Pietropaolo 2001, p. 137). Lang (1901, p. 157) fand übrigens außer Algen "die Reste von Insektenlarven; daneben grosse Nematoden (siehe Tafel-Fig. 3 und 4), welche oft grösser sind als der Längsdurchmesser des Blasenlumens, dann wieder ganze Schaaren von winzig kleinem Gewürm" in den Fallen von U. multifida (Schreibweise von Lang). 8. MN schreibt zum Thema der "falschen Tatsachen" außerdem (Hervorhebungen im Schriftbild von ihm): MN: "a.) Entgegen LÖNNIGs Behauptung verfügt Roridula nicht über die Fähigkeit, mittels Enzyme gefangene Insekten zu verdauen; vielmehr bedienen sich Blindwanzen [Pameridea] der willkommenen Nahrungsquelle und versorgen die Pflanze nur mittels ihrer Fäkalien mit Nährstoffen. Die von LÖNNIG erwähnten Phosphatasen sind aus mehreren im Text ausgeführten Gründen für eine Verdauung nutzlos." Was könnte man nun noch einem Autor entgegnen, der vorsätzlich die neuesten Befunde zu dieser Frage ignoriert, offenbar weil sie nicht zu seinen unrealistischen evolutionären Schemata passen? Ich hatte in der vorliegenden Arbeit die Beiträge von Plachno et al. von 2006 und 2009 zitiert: Plachno, B. J., Adamec, L., Lichtscheidel, I. K., Peroutka, M., Adlassnig, W. and J. Vrba (2006): Fluorescence labelling of phosphatase activity in digestive glands of carnivorous plants. Plant Biology 8, 813-820. Plachno, B. J., Adamec, L. and H. Huet (2009): Mineral nutrient uptake from prey and glandular phosphatase activity as a dual test of carnivory in semidesert plants with glandular leaves suspected of carnivory. Annals of Botany 104, 649-654. Dort legen die Autoren ihre Untersuchungsergebnisse vor, dass es sich bei Byblis und Roridula um echte Karnivoren ("true carnivores") mit "high activity of phosphatases" handelt, und zwar mit Nachweis von mineral uptake (N, P, K and Mg) von Drosophila prey bei Roridula auch ohne Symbionten. Zu den Phosphatasen schreiben Plachno et al. (2006, p. 814): "Hydrolysis of phosphate esters is a critical process of phosphorus metabolism at cellular, organism, and ecosystem levels. Phosphatases (phosphomonoesterases) represent a broad group of enzymes that catalyze the hydrolysis of phosphate esters (Feder, 1973). Acid phosphatases are common plant enzymes of low substrate specificity that appear to be important in the production, transport, and recycling of phosphorus (Duff et al., 1994). So far, a few authors (Clancy and Coffey, 1976; Robins and Juniper, 1980; Sirovä et al., 2003) have studied production of acid phosphatases by carnivorous plants. While proteases may not occur in some typical carnivorous plants (e.g., in Utricularia, Sirovä et al., 2003, or Byblis, Hartmeyer, 1997), we suggest the phosphatases as model digestive enzymes in this group. In our opinion, the phosphatases are indispensable for phosphate mobilization from prey carcasses and phosphate uptake may be essential for many carnivorous plants (e.g., Adamec, 1997)." Auf der Seite 815 stellen die Autoren unter anderem fest: "Plants cope with a deficiency of phosphorus in the soil by modifications of root morphology and by changes in phosphorus uptake and metabolism; in addition, the production of extracellular phosphatases can help to release inorganic phosphate from the environment (Chröst, 1991; Olczak, 1996; van Aarle et al., 2001; Nedoma et al., 2003; Strojsová et al., 2003; Hammond et al., 2004). Carnivorous plants are an example for the latter strategy since they use enzymatic exudates to digest phosphate compounds of their prey. Das entscheidende Ergebnis für unsere Fragestellung zu Byblis und Roridula lautet (p. 818): "In Byblis, only one author (Bruce, 1905, after Juniper et al, 1989) found evidence for the production of digestive enzymes, whereas Hartmeyer (1997) failed to detect proteases and, therefore, questioned the presence of carnivory in Byblis [aber 2005 gefunden]. Our results show that Byblis is able at least to digest phosphorus compounds. The total degradation of proteins, however, may depend on symbiotic organisms. So far, no digestive enzymes are known from Roridula. Prey degradation was thought to be performed only by symbiotic hemipterans (Ellis and Midgley, 1996; Anderson, 2005). In the leaf epidermis, however, we found phosphatase activity, but not in the glandular tentacles. Future research will show if the epidermis takes part in the utilization of prey, or if the phosphatases only reflect high metabolic activity in general. In ihrem Paper von 2009 stellen die Autoren nach Hinweis auf die Arbeiten von Anderson und Midgley (2003) und Anderson (2005), die bislang nur "'indirect' means of carnivory, mediated by commensal hemipteran bugs of the genus Pameridea" bei Roridula nachweisen konnten, fest (p. 649): Plachno et al. (2006) revealed phosphatase activity on the leaves of Roridula, suggesting also that direct carnivory may be possible." Ihre Versuche haben sie daraufhin mit Drosophila melanogaster durchgeführt (siehe die Details pp. 650/651) und zwar ohne commensal hemipteran bugs of the genus Pameridea. Ergebnis (2009, p. 651): "Application of model prey on the leaves revealed two distinct groups of plants based on uptake of mineral nutrients from prey (Table 3). Roridula and Drosophyllum were found to take up substantial amounts of both N (33-47% of the total N content), P (62-75%), K (44-86%) and Mg (33-39 %) from fruit flies. Except for N uptake in Drosophyllum, which showed considerable variability, uptake of all nutrients from flies was statistically significant in these two species. By contrast, usually very weak release of N (7-18 % of the total N content), P (1-3%), K (4-19%) and Mg (4-21 %) to flies, rather than nutrient uptake from flies, was found in the four remaining plant species, Proboscidea. Ibicella, Cleome and Hyoscyamus. However, this nutrient release to flies was not statistically significant. Das Ergebnis ist also völlig eindeutig: Ohne jegliche Beteiligung von Pameridea-Symbionten können Drosophyllum und Roridula "substantial amounts of both N (33-47% of the total N content), P (62-75%), K (44-86%) and Mg (33-39 %) from fruit flies aufnehmen, wobei die letztere jeweils höhere Zahl das Ergebnis für Roridula ist. "Despite the absence of phosphatase activity on Roridula emergences, the efficiency of P uptake and of other mineral nutrients in Roridula was very high (Plachno et al. 2009, p. 651). Steht damit nicht steht fest, dass die oben zitierte Auffassung (prey degradation was thought to be performed only by symbiotic hemipterans) falsch ist und MN mit einer weiteren falschen Tatsache (false fact) gearbeitet hat? Worauf beruht nun aber die prey degradation? Sind die extrazellulären Phosphatasen, die enzymatic exudates, daran beteiligt oder nicht? Hier werden die Autoren sehr vorsichtig, denn ihr bisheriger Versuchsaufbau kann diese Frage bedauerlicherweise nicht direkt und abschließend beantworten. Worin bestand die Versuchsanordnung? Antwort (p. 651): "Here, the applied fruit flies were evidently not in direct contact with foliar epidermis containing high phosphatase activity (Plachno et al., 2006) which would have enabled direct nutrient uptake through cuticular pores (Anderson, 2005). Wenn nun die Fruchtfliegen gar nicht in Kontakt mit der foliar epidermis containing high phosphatase activity waren wie ist dann degradation und uptake mit der geschilderten außergewöhnlich hohen Effizienz bei Roridula überhaupt möglich? Mit dem von den Autoren des Weiteren unterbreiteten Vorschlag, dass nun anstatt der Phosphatasen "autolysis of prey tissues and release of digestive enzymes by microbial commensals" (p. 653) die entscheidende Rolle spielen könnten ("may occur") (aber keinesfalls die ohnehin nicht an den Versuchen beteiligten Pameridea-Symbionten), bleiben allerdings noch mehrere Fragen unbeantwortet. Sehen wir uns diese Hypothese einmal etwas näher an: Selbst nach Autolyse der Drosophila-Gewebe und dem Abbau durch Mikrobenenzyme muss ja irgendwann die phosphatasenreiche (Stichwort: enzymatic exudates), durch die speziellen cuticular gaps and pores (Anderson) ausgezeichnete und somit durchlässige unten liegende, großflächige Epidermis erreicht werden. Andernfalls würde praktisch nichts von Drosophila absorbiert werden. Und wenn nicht die gesamte Zersetzungsarbeit schon vorher getan wurde (was denkbar unwahrscheinlich ist; siehe dazu die Abbildungen aus M. Reiner und anderen unten), spielen logischerweise auch die extrazellulären Phosphatasen unausweichlich eine bestimmte Rolle in dem Geschehen. Plachno et al. aber konnten das nicht feststellen, da sie ausnahmslos mit schon toten Fliegen arbeiteten, die nur mit den resin producing tentacles, aber nicht mit den enzymatic exudates der Epidermis in Kontakt gekommen sind. Oben links: Roridula gorgonias mit zahlreichen gefangenen Insekten (vor allem Fliegen, aber im Bild unten auch eine Wespe, aber hier ohne Pameridea). In vielen Fällen kommen die Tiere auch direkt in Kontakt mit den phosphatasenreichen Exsudaten der Epidermisoberfläche (aus Martin Reiner 2004: Roridula & Pameridea die Taupflanze und ihre Wanzen: http://www.drosophyllum.com/Bilder/Gross/Roridulagorgonias05g.jpg .) Siehe dazu weiter die Frage nach Schmerzempfinden bei Insekten unter http://www.weloennig.de/JoachimVetter.pdf (dennoch tun mir die Tiere leid). Und ein weiterer Aspekt. Anderson bemerkt zum Thema Absorptionsvermögen und -geschwindigkeit der Epidermis sowie der Inkorporation in das Blattgewebe der Roridula-Pflanzen (2005, p. 760): "The extensive staining by neutral red in Roridula suggests that the majority of epidermal cells are highly absorbtive and that the entire epidermis has an absorptive function [im Gegensatz etwa zu Drosophyllum, die "only a few specialized digestive cells nach Joel und Juniper aufzuweisen hat]. These results lend credence to the results of Ellis and Midgley (1996) who show that nitrogen from trapped flies is very rapidly absorbed by Roridula plants. Their results show that substantial amounts of fly nitrogen are incorporated into plant leaves after only 72 h of capture. They postulate that such rapid nitrogen incorporation can only take place if digestion is immediate. Da weder Ellis und Midgley (1996) noch Anderson (2005) etwas von den epidermalen enzymatic exudates samt extrazellulären Phosphatasen wussten, schlossen sie direct carnivory aus und konzentrierten sich auf die Stoffwechsel-Endprodukte (faeces) der Hemipteren (Pameridea-Arten). Plachno et al. (2009) aber haben nachgewiesen, dass bei Roridula auch die direkte Karnivorie möglich ist, und zwar mit dem erstaunlichen Ergebnis, dass um diesen Kernpunkt noch einmal zu betonen 47% N, 75% P, 86% K und 39% Mg aus den Fruchtfliegen ohne die geringste Beteiligung von Pameridea roridulae aufgenommen werden.
Links und rechts: Fliege in direktem Kontakt mit der enzymatic-exudates-reichen Roridula-Epidermis und dazu ein Exemplar von Pameridea roridulae. Ergebnis: Effiziente Zusammenarbeit von direkter und indirekter Karnivorie. Aus http://www.fischermans-antik.de/ShowTemplate-Dateien/Eigene%20Bilder%20Fleischis/CK17/60.JPG und http://www.cpukforum.com/forum/index.php?showtopic=22402 . Im Übrigen ist das Phänomen des Mutualismus zwischen Roridula und Pameridea selbst ein schweres Problem für die Synthetische Evolutionstheorie, ebenso mit der Spinne Synaema marlothii ein Punkt der eine eigene Abhandlung wert wäre (vgl. zur morphologisch-anatomisch und physiologisch präzisen Abstimmung (Tier- und Pflanzenreich übergreifende Synorganisation der Arten aufeinander) z. B. http://www.drosophyllum.com/deutsch/roridula.htm Die Aussage von p. 2 oben ist damit für Roridula noch einmal im Detail bestätigt: Die Gattungen Byblis und Roridula sind weder direkte noch 'funktionale' Vorstufen zu den Lentibulariaceae. Es handelt sich bei diesen beiden Gattungen vielmehr um echte Karnivoren ("true carnivores") mit "high activity of phosphatases" mit Nachweis von mineral uptake (N, P, K and Mg) von Drosophila prey bei Roridula ohne Symbionten (vgl. p. 103: Plachno et al. 2006, 2009; Byblis: "Verdauungsenzyme" auch nach Barthlott et al. 2004), deren Drüsen anatomisch eine Sondergestaltung aufweisen. Dass Roridula weder eine direkte oder auch nur funktionale Vorstufe zu den Lentibulariaceen ist, zeigt sich weiter in der Tatsache, dass die resin producing tentacles keine Vorstufe zu den mucilage producing tentacles sind: the glue of Drosera, Drosophyllum, Triphyophyllum, Pinguicula, Byblis is mucilage (water-based with high sugar content. It has to be water based in order to dissolve enzymes in). In contrast, the glue of Roridula is a resin. It never evaporates and cannot be washed away by rain. But big disadavantage: no enzymes can be dissolved in the glue droplets of Roridula [was unter anderem erklärt, warum Plachno et al. 2006 und 2009 dort keinerlei Phosphatasen nachweisen konnten]) - Andreas Fleischmann 2010: http://www.cpukforum.com/forum/index.php?showtopic=35105 . Eine gute Zusammenfassung und Erklärung der Methoden und Befunde sowie der Bedeutung der Arbeiten von Plachno et al. (2006 und 2009), speziell zu den Phosphatasen, findet der daran interessierte Leser bei John Brittnacher von der International Carnivorous Plant Society http://www.carnivorousplants.org/cp/Digestion.php (Zugriff 28. 3. 2011: "It is very clear from the data that phosphatases are important to carnivores). Der Leser beurteile bitte wieder selbst, ob meine Einschätzung, dass MNs Tatsachenbehauptung zu Roridula (als Vorstufe zu den Lentibulariaceen; vgl. hier Punkt 88, p. 106) sowie MN 2010, p. 14: "Roridula nimmt lediglich die Nährstoffe aus den Fäkalien ihrer Symbiosewanzen (Pameridea) über die Spaltöffnungen der Blätter auf" oder Roridula, "die nachgewiesenermaßen keine Tiere verdauen kann" (p. 11) nach dem vorliegenden Tatsachenmaterial zu den false facts gehört oder nicht. 9. Zu Byblis und noch einmal Roridula (Hervorhebung im Schriftbild von MN): MN: "b.) Byblis ist nach Ansicht einiger Karnivoren-Spezialisten durchaus ein nächster Verwandter von Pinguicula (genauer: der Wasserschlauchgewächse)." " nach Ansicht einiger Karnivoren-Spezialisten " die anderen sind jedoch gegenteiliger Auffassung! (Vgl. die Details p. 104 und Brittnacher zitiert unten). Die, wie auch im vorliegenden Falle, oft erstaunlich widersprüchlichen Rekonstruktionen phylogenetischer Verwandtschaftsverhältnisse auch zu Byblis zeigt einmal mehr, dass man diese nicht mit der Sicherheit der DNA-Vaterschaftstests gleichsetzen kann. Das führt uns vielleicht noch einmal zur Frage, inwieweit MN die gravierenden Unterschiede zwischen der Sicherheit des Vaterschaftstests (zur 99,9% sicheren Identifikation eines Kandidaten aus einer ausgewählten Gruppe innerhalb der Art) und der Unsicherheit phylogenetischer Rekonstruktionen (Makroevolution) verstanden hat, deren widersprüchliche Ergebnisse er (wenn auch unbeabsichtigt) mit der obigen Aussage zu Byblis bestätigt. Zur Behauptung Byblis sei eine funktionale Vorstufe zu den Wasserschlauchgewächsen vgl. weiter die Argumentation und die Fakten in der vorliegenden Arbeit zum Punkt 81, p. 104 (wie: "Differences in gland morphology support parallel acquisition of carnivorous traits in Byblis (Byblidaceae), Ibicella (Martyniaceae), and Lentibulariaceae. Digestive glands in all three genera of Lentibulariaceae are attached to vessels, unlike secretory glands of Byblidaceae and Martyniaceae that rest on at least two epidermal cells" Müller; auf diese Sondergestaltung, auf die unterschiedliche Anatomie, hätte MN in einer überzeugenden Diskussion eingehen müssen statt sich widersprechende Autoritäten zu zitieren). Zum Punkt 79, p. 102 womit wir auch wieder zu Roridula zurückkommen (MN: "Klebrige Drüsenhaare zur Abwehr von Schadinsekten finden sich schon bei nichtkarnivoren Pflanzen, etwa bei Roridula. Diese Pflanze hält die Insekten mittels eines klebrigen Sekrets fest.") und Punkt 80, pp. 103/104 (MN: "Der daran anknüpfende Evolutionsschritt bestand ganz offensichtlich in der Differenzierung der Drüsenhaare, wobei einige Drüsenhaare zusätzlich Verdauungssekrete absondern, um die festsitzenden Insekten zu zersetzen (HEUBL et al. 2006, 827, Fig. 6).") sowie Punkt 81, p. 104 (MN: "Byblis produziert wie Roridula ein klebriges Sekret, ") siehe die oben zitierten Fakten: Es handelt sich um völlig unterschiedliche Sekrete: water-based mucilage to dissolve enzymes bei Byblis vs. resin bei Roridula " no enzymes can be dissolved in the glue droplets of Roridula." Oder B. Rice 2005/2009: "In the early 1900s, the botanist Marloth pointed out that, unlike other sticky flypaper plants that produce leaves covered by mucus glands, the leaves of Roridula are covered with resin glands. This is a key difference. Mucus is water-based, resin is not. (Did you ever try to wash pine sap off your hands just using water?) Digestive enzymes and nutrients can easily diffuse through mucus, but they cannot diffuse through resin (vgl. http://www.sarracenia.com/faq/faq5500.html ). ("In the early 1900s": Zur Richtigkeit und Bedeutung älterer Arbeiten siehe die Ausführungen oben.) Resin glands in Roridula: MN hat das inzwischen auch festgestellt (2010), erwähnt aber nicht, dass damit Roridula als funktionale Vorstufe und als Beispiel für seine früheren Aussage ("wobei einige Drüsenhaare zusätzlich Verdauungssekrete absondern"), verfehlt ist von seiner bisherigen Weigerung, die Forschungsergebnisse von Plachno et al. (2006, 2009) zur Roridula-Epidermis (repetitio est mater studiorem: enzymatic exudates samt extrazellulären Phosphatasen und uptake von 47% N, 75% P, 86% K und 39% Mg aus Fruchtfliegen ohne die geringste Beteiligung von Pameridea roridulae) zur Kenntnis zu nehmen, einmal ganz abgesehen. Ein Kernpunkt zur Unterscheidung von karnivoren und nichtkarnivoren Pflanzen ist in diesem Zusammenhang weiter die folgende Feststellung von Plachno et al. (2009, p. 651): "True carnivorous plants possess not only anatomical modifications to trap animals and external enzyme activily in traps, but crucially show sufficient mineral nutrient absorption from prey carcasses to support plant growth and fitness in competition with non-carnivorous plants (Juniper et al.. 1989: Adamec, 1997 [siehe zum Thema "fitness in competition " die Ausführungen oben]). Of all six species tested 39 only Roridula and Drosophyllum have been found to take up nutrients (N, P, K. Mg) from applied fruit flies (Table 3: cf. Adamec, 2002). Roridula ist nach diesen Befunden eindeutig eine karnivore Gattung und sie ist durch ihre Spezialisierungen (resin glands zum Insektenfang in Zusammenarbeit mit struktureller und funktionaler Sondergestaltung der Epidermis mit sufficient mineral nutrient absorption from prey carcasses) weder eine direkte noch eine funktionale Vorstufe zu anderen Karnivoren oder zu den Karnivoren überhaupt. Und dass die ohnehin zu den echten Karnivoren zählende Byblis ebenfalls Spezialisierungen (Heterobathmien, specialization-crossings, chevauchements de specialisation) aufweist, die sie auch nur als "funktionelles" Bindeglied zu den Lentibulariaceen unbrauchbar machen, ist den meisten jüngeren und älteren Beschreibungen leicht zu entnehmen. Beispiele: D'Amato 1998, p. 181: "The leaves are fine and linear, radiating in all directions from the stem." J. and P. Pietropaolo 2001, p. 105: "In cross-section the base of the leaf is triangular with rounded edges but becomes almost circular in cross-section at the tip." Barthlott et al. 2004, p. 84: "Die Blätter aller Arten sind in der Knospenlage nach außen aufgerollt; diese Form der Knospenlage ist äußerst selten und kommt auch bei Drosophyllum lusitanicum vor." Slack 2000, pp. 95/96: "When mature they [the leaves] are held at an acute angle to the stem; the underside rather then the top surface of the leaf is therefore the most exposed, and provides the most obvious surface on which an insect in flight might land. Not surprisingly, then it is the underside of the leaf which is used principally by the plant to detain its prey. This is heavily scattered with stalked, mucilage-secreting glands, but these are almost entirely lacking on the upper surface, where those that occur are of diminutive size." [Slacks Aussage zum Winkel scheint in erster Linie auf junge Blätter zuzutreffen. Siehe weiter die differenzierte Diskussion zu Byblis von Lloyd 1942, pp. 95-96.] Und noch einmal Slack 2000, p. 96: "While the stalked retentive glands are large and very conspicuous, the digestive glands, though extremely numerous, are of microscopic size and quite invisible to the eye [der Ausgangspunkt sollten vielmehr die large and very conspicuous glands gewesen sein, die dann später zusätzlich Verdauungsenzyme produzierten]. They are arranged in single rows, and are sunk for protection in little furrows. They are to be found not only on both sides of the leaf, but also on the stems. Brittnacher fasst die bisherigen Untersuchungsergebnisse zum Thema Byblis und Evolution wie folgt zusammen ( http://www.carnivorousplants.org/cp/EvolutionLamiales.php Zugriff am 29. 3. 2011): "There is not much to say about the evolution of Byblis at this point. In the DNA phylogenies it tends to fall in with families of plants that used to be considered part of the Scrophulariaceae and not close to the other carnivores. It has a unique design for its glands and appears to have evolved carnivory independently of the the other carnivores. Zu Detailunterschieden in der Struktur und Zytochemie zwischen den digestive glands von Byblis und Pinguicula vgl. z. B. die Abbildungen in http://www.carnivorousplants.org/cp/EvolutionLamiales.php nach Müller et al. 2002, dort genauso für die mucilage glands; siehe weiter auch die Arbeiten von Lloyd 1942, Y. Heslop-Harrison und J. Heslop-Harrison 1980, 1981, Vassilyev und Muravnik 1988, Müller et al. 2002, 2006, Plachno 2007). Byblis besitzt "als einzige Karnivore Verdauungsdrüsen, die direkt am Stamm und an den Fangarmen sitzen" http://www.fleischfressendepflanzen.de/db/gattung.ffp?id=6 . Auch der "Klebstoff" scheint sich in Byblis und Pinguicula zu unterscheiden ( http://www.cpukforum.com/forum/index.php?showtopic=31967 2009, er arbeitet möglicherweise mit einem Betäubungsmittel). Berücksichtigt man dazu die auf den Seiten 103/104 aufgeführten Fakten, so wird deutlich, dass auch die Gattung Byblis weder direkt noch funktionell als eine Vorstufe zur Blattfalle von Pinguicula aufzufassen ist. Es handelt sich selbst nach evolutionstheoretischen Voraussetzungen vielmehr um eine unabhängig (konvergent) entstandene, ganz eigengeartete Gattung mit entsprechendem Sonderstatus was auch auf die schon oben behandelten Gattungen Roridula und Drosophyllum zutrifft. Slack zählt die Gattungen Byblis und Drosophyllum passend zu den "passiven Klebfallen (2000, p. 95, Kapitel 6: "The Passive Flypapers, wie schon Lloyd 1942) und schreibt einleitend: "The plants in this group belong to two genera: Byblis and Drosophyllum. There is certain superficial resemblance between the two in their general appearance and in various features which they share in common. However, this is purely an accident in evolution, for they belong to quite separate families. Each plant secures its prey by means of a sticky mucilaginous substance secreted by numerous stalked glands. Both also possess other, sessile glands, which secrete enzymes around the victims' bodies and are digestive in function. In this way all but the chitinous husks and wings of the victims are broken down and dissolved into a fluid which may thus be readily assimilated by the plant. These plants are described as 'passive' in the sense that they have not evolved that power of movement which we shall see in the sticky leaves of the Butterworts, and in the gland-tipped 'tentacles' and often in the leaves themselves of the Sundews. But it is very questionable whether such added sophistications would be of any real advantage to them at all, for both genera are so designed that they can efficiently catch and digest a truly remarkable number of insects. Mit dieser Einschätzung von Slack sind wir wieder beim oben schon ausführlich behandelten Thema der äußerst fragwürdigen selektionstheoretisch notwendigen Weiterentwicklung der bestehenden Formen und Gattungen (die sich überdies paläontologisch häufig als living fossils geoutet haben) und können derzeit nur feststellen, dass Gattungen wie Byblis auch selektionstheoretisch weder direkt noch funktionell eine Vorstufe zu Pinguicula oder überhaupt zu den Lentibulariaceen sind. Und wieso in aller Welt sollten sich die hypothetischen Vorläufergattungen und -Arten grundsätzlich anders verhalten haben? Vielleicht weil es sonst die hypothetische (Makro-)Evolution nicht gäbe? Wie steht es jedoch mit einigen "protokarnivoren" Spezies (Spomer 1999, p. 99, von mir nach Pflanzenfamilien geordnet) wie: Cerastium arvense (Caryophyllaceae) Zunächst sei kurz erwähnt, dass der Begriff der "Protokarnivorie nicht unumstritten ist: "Some authors prefer the term "protocarnivorous" because it implies that these plants are on the evolutionary path to true carnivory, while others oppose the term for the same reason. The same problem arises with "subcarnivorous." Donald Schnell, author of the book Carnivorous Plants of the United States and Canada, prefers the term "paracarnivorous" for a less rigid definition of carnivory that can include many of the possible carnivorous plants ( http://en.wikipedia.org/wiki/Protocarnivorous_plant ). Nach Spomer (1999) is Proteaseaktivität auf den Blättern der oben genannten Spezies nachgewiesen. Macht das Nelken, Rosen und Kartoffeln zu (proto-) karnivoren Pflanzen? "These species tested positive for protease activity, though it is unclear whether the protease is produced by the plant or by surface microbes. 40 "The source of proteinase activity, whether plant or surface microbes, is yet to be investigated (Spomer 1999, p. 101). Aber Geranium viscossisimum und Potentilla arguta "absorbed digestion products of a protein applied to the leaves" (nach Plachno et al. 2009, p. 651; und Spomer selbst 1999, p. 98: " the breakdown products were absorbed and translocated). Dabei handelte es sich um Bruchstücke von 14C-markiertem Protein der Alge Synechococcus leopoliensis. Zusätzlich kann man noch Stylidium erwähnen (Darnowski et al. 2006). Zur Letzteren ist jedoch die Frage nach nutrient uptake noch offen. 41 Jetzt wollen wir die Situation einmal hypothetisch stark vereinfachen und annehmen, dass alle oben genannten Arten in der Lage sind, zumindest in geringem Maße "digestion products of a protein applied to the leaves" absorbieren können 42 . Würde diese Fähigkeit die oben genannten und vielleicht auch noch die zahlreichen weiteren Pflanzenarten mit sticky leaves zu karnivoren Pflanzen machen? Oder zumindest zu Protokarnivoren? Die Liste dieser sticky-leaves-Pflanzenarten ist sehr lang hier nur einige wenige der zahlreichen "glutinosa"-Beispiele, die sich durch das ganze Alphabet ziehen: Alnus glutinosa, Arctostaphylos glutinosa, Baccharis glutinosa, Calytrix glutinosa, Cuphea glutinosa, Eragrostis glutinosa, Eremophila glutinosa, Erica glutinosa, Eriodictyon glutinosa, Eucryphia glutinosa, Gutierrezia texana var. glutinosa, Lysimachia glutinosa, Nepeta glutinosa, Olearia glutinosa, Primula glutinosa, Rehmannia glutinosa, Sida glutinosa, Salvia glutinosa, Triantha glutinosa, Tofieldia glutinosa. Würden sticky leaves und die Möglichkeit der Absorption von zumindest kleineren Molekülen nicht schon fast das halbe Pflanzenreich zu "Protokarnivoren" machen? Dass die Sache jedoch nicht so einfach liegt, zeigen unter anderem die Untersuchungen zur möglichen Protokarnivorie der sticky-leaf-Arten Lathraea squamaria, Salvia glutinosa und Rubus phoenicolasius von S. Pohl 2009: "Aufgrund der Ergebnisse dieser Arbeit ist keine der drei Pflanzen als protokarnivor zu bezeichnen, da sie offenbar keinen Nutzen von den gefangenen Tieren haben." (Vgl. http://othes.univie.ac.at/6995/ .) Wenn es nun evolutionär einfach und vorteilhaft wäre, allein auf der Basis von sticky leaves und selektiver Absorption von Molekülen Karnivoren wie Utricularia, Aldrovanda oder Dionaea mit ihren perfekten Fangmechanismen zu entwickeln etc. warum hat das dann in dem gewaltigen Zeitraum von rund 1 Milliarde Jahren und unzähligen von Mutationen in Billionen von Pflanzen nur so selten geklappt? Ich erinnere noch einmal an die Feststellung von Juniper, Robins and Joel: " there are very few carnivorous plants. There is none, with the exception of a few fungi, below the angiosperms; there are no herbs, no shrubs or trees and there are large areas of the phylogenetic table (see Fig. 1.2) where carnivory does not seem to have evolved at all. Ähnlich Benzinq (1987, p. 364): "But despite its obvious advantages and substantial geological history, botanical carnivory remains a minor nutritional mode, apparently because prey use is usually not the most economical way for plants to secure nutrients. Worin liegt jetzt aber der Unterschied zwischen den normalen sticky-leaves-Pflanzen und etwa Roridula und Byblis? M. Honda hat (2010) zu den Karnivoren folgenden Definitionsvorschlag unterbreitet ( http://www.inet1000.com/IPW_BOOK/Text.htm ): "The exact definition of carnivorous plants may differ among authors. But to be included in this exclusive society of plant carnivores, a plant must minimally exhibit the ability to capture prey, digest it, and absorb its nutrients [in substantial amounts]. Many carnivorous plants also provide various attractions to lure prey. Last but not least, a/the plant must derive survival benefit from carnivory. The general description of carnivorous plants then becomes: 1 Attraction of prey Roridula und Byblis erfüllen alle diese Bedingungen vollendet (carnivorous syndrome). Oben hatten wir schon den Gegensatz zwischen Roridula und Drosophyllum (uptake of substantial amounts of N, P, K, and Mg from Drosophila prey) auf der einen Seite und sticky-leaves-Spezies von Proboscidea, Ibicella, Cleome und Hyoscyamus (no uptake) auf der anderen zitiert. Die bisherigen Daten legen die Schlussfolgerung nahe, dass bei Roridula, Drosophyllum und Byblis ein deutlicher quantitativer und qualitativer Sprung im Vergleich zu den nichtkarnivoren Arten vorliegt. Es gibt zwar zahlreiche z. T. widersprüchliche, aber innerhalb ihres jeweiligen Rahmens manchmal recht plausible Spekulationen, wie sich die Karnivorie im Pflanzenreich entwickelt haben könnte, aber "plausible stories need not be true" (Gould): Tatsache ist vielmehr: "There is no clear picture of how often, by what route, or when carnivory emerged during plant evolution" (Benzinq). 44 Innerhalb des nach Honda und weiteren Autoren (siehe Fußnote) zitierten Definitionsrahmens gibt es nun zahlreiche unterschiedliche Attraktions- und Fangmethoden, digestion of prey etc. in unterschiedlichen Komplexitätsstufen (wie oben ausführlich diskutiert), die sogar am gleichen Standort zusammen mit karnivoren (nahe und entfernt verwandten) und nichtkarnivoren Arten (von Algen bis zu den Angiospermen) erfolgreich auftreten und damit selektionstheoretisch gleichwertig sind. Siehe zur Ergänzung auch die langen Listen der zusammen mit einigen Pinguicula-Arten (Pinguicula vulgaris, P. grandiflora und P. lusitanica) vorkommenden nichtkarnivoren Spezies, die Yolande Heslop-Harrison (2004) zusammengestellt und publiziert hat: vgl. dort die Tables 2, 5, and 8 http://onlinelibrary.wiley.com/doi/10.1111/j.0022-0477.2004.00942.x/pdf . Eine Detaildiskussion, warum die oben aufgeführten Nelken, Rosen, Primeln, Kartoffeln etc. mit ihren sticky leaves nicht zu den (proto-)karnivoren Pflanzen gehören, scheint mir derzeit nicht notwendig zu sein. Einige Hauptpunkte hat S. Pohl anhand der von ihr untersuchten sticky-leaves-Arten schon herausgearbeitet (siehe oben) und der an weiteren Punkten interessierte Leser wende bitte die obige Definition von Honda (inklusive der Ergänzungen in der Fußnote) auf diese Frage weiter an. 10. Zu den unterschiedlichen Differenzierungs- und Komplexitätsstufen im Organismenreich generell nicht nur bei den Karnivoren vielleicht folgende simple Veranschaulichungen aus der Technik (selbstverständlich hinken Beispiele fast immer, wer aber die logische Struktur der folgenden Beispiele versteht, wird keine Schwierigkeiten haben, sie auf die unterschiedlichen Differenzierungsstufen der Karnivoren anzuwenden): Beweist die Tatsache, dass es (Kinder-)Roller, kleine und große Fahrräder, Dreiräder, Kinderwagen ('Vierräder'), Motorräder, Autos (PKWs und Lastwagen/Trucks mit mehr als 4 Rädern) etc. gibt, dass sich alles vom Roller ableitet und überdies von selbst entstanden ist (Selbstorganisation)? Und dass etwa bei einem Mercedes 240 E ein funktional irreduzibles core system nicht vorhanden wäre? Zeigt uns die Serie Hundehütte, kleine Laube, Lehmhütte, Einfamilienhaus, Villa mit Swimmingpool, Schloss Neuschwanstein, Buckingham Palace, dass sich die Schlösser allesamt von der Hundehütte ableiten und ohne Geist, Plan und Ziel entstanden sind? Siehe auch die Veranschaulichungen auf der Seite 107 oben und Casey Luskin 2006 45 . Oder vielleicht mit MN (etwas verändert): "Arten wie Roller, Dreiräder, Fahrräder und Motorräder belegen auf eindrucksvolle Weise, dass auch komplex(er) gebaute Maschinen wie ein Mercedes 240 E keineswegs durch eine gezielte (Syn-)Organisation aller für die Funktion wesentlichen Komponenten entstehen müssen." Zu biologischen Beispielen vgl. Auge widerlegt Zufalls-Evolution 46 und Die Evolution der Langhalsgiraffe (Giraffa camelopardalis) was wissen wir tatsächlich? Zwei Teile 47 ; siehe zur Problematik der oft einfach "als gegeben" vorausgesetzten einzelnen Schritte bei den Karnivoren auch die Anmerkungen auf den Seiten 167/168 der vorliegenden Arbeit. Gibt es nun Protokarnivoren? Ja, und zwar genau so sicher wie ein Kinderroller oder ein Fahrrad ein Protomercedes ist. Unter evolutionstheoretischen Voraussetzungen bleibt nur wieder völlig unerwartet, dass sich die vielen Arten von 'Protokarnivoren' ("...the term "protocarnivorous"... implies that these plants are on the evolutionary path to true carnivory...") auch in Millionen von Jahren nicht weiterentwickeln konnten. 11. MN: "c.) Die Behauptung, "Heliamphora tatei [sei] primitiv und [produziere] keine Enzyme", hat LÖNNIG erfunden und mir wahrheitswidrig untergeschoben." [Fett von MN.] Hier noch einmal MNs hypothetische Serie funktional zunehmender Komplexität (vgl. Punkt 88, p. 106): Roridula
gorgonias Von Sarracenia purpurea hatte der Autor zuvor ohne Einschränkung behauptet, dass diese Spezies keine Verdauungsenzyme produziere (wörtlich "die keine Verdauungssäfte herstellt", vgl. hier Punkt 63, p. 73). Unter Punkt 64 (p. 75) bezeichnet er "die Sumpfkrüge (Heliamphora)" als Beispiel für rezente "passive Fallgruben (Gleitfallen)". Passiv heißt hier unter anderem, dass sie ebenfalls keine Enzyme produzieren (und zwar wiederum ohne jegliche Einschränkung von MN) und er nennt dann später ausdrücklich Heliamphora tatei als Beispiel für seine konstruktive Reihe vor dem nächsten Schritt Sarracenia pupurea (p. 106). Heliamphora tatei ist somit nach MNs Schema primitiver als Sarracenia purpurea. Schlussfolgerung: Keine von beiden Arten produziert nach MNs Darstellung irgendwelche Enzyme. Was an den Vertretern von Heliamphora sonst noch primitiv sei, sagt er uns unter Punkt 65, p. 82: "Bei diesen Vertretern kann man noch genau erkennen, wie sich ein normales Laubblatt zu einem Schlauchblatt umformte (Abb. 46)." In der speziellen Zusammenfassung zu den Karnivoren habe ich Folgendes festgestellt (p. 2): An Roridula gorgonias und Byblis gigantea schließt MN "konstruktiv" Heliamphora tatei an (siehe Punkt 64, p. 75 und expressis verbis Punkt 88, p. 106 unten) und arbeitet dabei mit den "false facts", dass Heliamphora tatei primitiv sei und keine Enzyme produziere. Beides ist nachweislich falsch. In der Klammer hatte ich zunächst nur geschrieben ("siehe u. a. Punkt (88) unten") und das Weitere später ergänzt ("siehe Punkt 64, p. 75 und expressis verbis Punkt 88, p. 106 unten"). Der Leser entscheide bitte angesichts dieser Tatsachen wieder selbst, wer hier etwas Falsches ("false facts") erfunden hat und wer wem etwas 'wahrheitswidrig unterschiebt'. 12. MN: "d.) Sarracenia purpurea produziert zwar, entgegen ursprünglichen Erkenntnissen, im ersten Jahr zeitweilig eigene Hydrolasen. Ab dem 2. Jahr wird die Verdauung allerdings von den Destruenten (z. B. Bakterien) übernommen, welche die Falle besiedeln." [Fett von MN.] Wie schon erwähnt, behauptet MN in seinem Originalbeitrag (2009; hier unter Punkt 63, p. 73 wiedergegeben) ohne jegliche Einschränkung, dass Sarracenia purpurea keine Enzyme produziere (das sollte eine Ausnahme bei den Sarraceniaceen sein. MN: "Eine Ausnahme ist S. purpurea, die keine Verdauungssäfte herstellt. Dort übernehmen Bakterien (!) die Verdauung."). Ich hatte diesen Punkt in der Zusammenfassung wie folgt beanstandet: Der Autor arbeitet weiter mit "false facts" indem er die Reihe mit Sarracenia purpurea und Utricularia multifida fortsetzt und im deutlichen Gegensatz zu den Tatsachen behauptet, Sarracenia purpurea würde ebenfalls keine Verdauungsenzyme produzieren Es ist nun wirklich eine falsche Tatsache, dass Sarracenia purpurea keine Verdauungsenzyme produziert. Der nicht ganz koschere Rechtfertigungsversuch ("Sarracenia purpurea produziert zwar, entgegen ursprünglichen Erkenntnissen, im ersten Jahr zeitweilig eigene Hydrolasen.") hingt gleich an mehreren Stellen: Dass S. pupurea Verdauungsenzyme produziert, war schon 12 Jahre vor MNs Beitrag (2009) bekannt (siehe die Details hier p. 74) und die "ursprünglichen Erkenntnisse" waren keine "Erkenntnisse", sondern Missverständnisse aufgrund mangelhafter Forschung und waren seit rund 12 Jahren überholt (er hätte das also bei einer sorgfältigen Recherche vorher wissen können). MN: " im ersten Jahr zeitweilig Hydrolasen": Bei den Hydrolasen handelt es sich um Proteasen, RNasen, Nukleasen und Phosphatasen. Chitinasen wurden nicht nachgewiesen (aber weiter Enzyme können nicht ausgeschlossen werden). Was das "zeitweilig" alles bedeutet, habe ich auf der Seite 74 weiter ausgeführt (Länge und Induzierbarkeit; weitere Details zum Thema insbesondere zu den Hydrolasen bei Gallie und Chang 1997: http://www.plantphysiol.org/content/115/4/1461.full.pdf+html ). Noch weiter zur Aussage " im ersten Jahr zeitweilig": "Even though a trap can live in excess of 1 year, the bulk of prey is caught within the first 50 days after a trap opens" (Gallie and Chang 1997, p. 1461), Induzierbarkeit auch noch nach 4 Monaten. "Digestion in the pitcher is aided, especially during the second year, by the commensal organisms that live in the pitchers (Barry Rice 2005/2010 http://www.p.com/faq/faq5538.html , man beachte besonders die vorsichtige Formulierung "aided, nicht "substituted). Auf eine Anfrage von MN behauptete Rice am 11. 4. 2010 jedoch plötzlich das Gegenteil dessen, was ich gerade von ihm 2005/2010 zitiert habe. Ich fragte B. Rice (am 2. und 3. 4. 2011), ob und falls ja, von wem diese Frage wissenschaftlich untersucht worden ist. Antwort: Seines Wissens gibt es nur die Untersuchungen von Gallie und Chang (ganz in Übereinstimmung mit meinen eigenen Recherchen; auch Brunners Aussage, dass eigene Enzyme im zweitem Jahr fehlen, ist nur eine Vermutung), d. h. niemand hat diese Frage für das zweite Jahr bisher untersucht. Ergo: Die Frage ist noch offen. Kurz: Anstatt seinen Fehler, dass S. pupurea keine Verdauungsenzyme produziere, unumwunden zuzugeben, versucht er alles Mögliche 48 , um seine falsche Aussage so klein wie möglich zu reden und den Anschein zu erwecken, dass er manche Dinge kaum wissen konnte, bzw. dass er für das zweite Jahr ja doch recht hatte 49 . Kurz: MN hat genau passend für seine evolutionären Erklärungs- und Rechtfertigungsversuche ungeprüft eine spätestens seit 1997 überholte Aussage übernommen und uneingeschränkt behauptet, S. purpurea produziere keine Verdauungsenzyme. Diese Behauptung hat er als Tatsache ausgegeben folglich hat er mit einer falschen Tatsache operiert. Und jetzt übernimmt er (wieder ungeprüft und scheinbar passend für seine Evolutionsvorstellungen und Rechtfertigungsversuche) die Behauptung, dass vom zweiten Jahr an die Verdauung in den pitchers (ausschließlich) von Destruenten übernommen werde (d. h. dass S. pupurea im zweiten Jahr keinerlei Verdauungsenzyme mehr produziere), obwohl niemand diese Frage bisher wissenschaftlich genau untersucht hat und überdies diese Behauptung selbst aus selektionstheoretischer Sicht nicht gerade wahrscheinlich ist: Denn für die Selektionstheorie, die MN mit seinen Ausführungen verteidigen möchte, erhebt sich die Frage: Welchen Selektionsvorteil sollte denn der völlige Verlust der präzis induzierbaren Produktion von digestive enzymes im zweiten Jahr haben? Und wenn es ohne Verdauungsenzyme genauso gut geht, wozu produziert S. purpurea dann im ersten Jahr überhaupt welche? Forschungsvorschlag: Können die pitchers von S. pupurea auch im zweiten Jahr noch Verdauungsenzyme induziert produzieren? Falls ja was nicht unwahrscheinlich ist welche? Fall nein wie ist dann der Verlust selektionstheoretisch zu erklären (welchen Vorteil hat der völlige Verlust)? Zu meinen Forschungsvorschlägen möchte ich hier kurz ergänzen, dass in einem Pilotversuch mit Pinguicula zur Untersuchung der Hypothese Barthlotts et al. ("Durch Herabsenken der Fangblätter ins Erdreich kann man sich die Entstehung der Genlisea-Reusenfallen und Utricularia-Fangblasen vorstellen") genau das eingetroffen ist, was von meiner Seite biologisch begründet prognostiziert wurde: Schon nach einigen Wochen war von den ins Erdreich herabgesenkten Fangblättern von Pinguicula praktisch nichts mehr übrig 50 (sogar zu meiner eigenen Überraschung, dass das derart schnell ging). "Ende der Weiterentwicklung!" (Vgl. p. 90 oben.) 13. Wie sicher ist eine wissenschaftliche Theorie und Position, die es immer wieder nötig hat zu ihrer Verteidigung von einer weitverbreiteten Ablehnung, Unwissenheit und Vorurteilen im Publikum auf völlig anderen Gebieten profitieren zu wollen etwa zur Religionszugehörigkeit von Wissenschaftlern (vgl. pp. 9-15 von http://www.weloennig.de/Die_Affaere1.pdf )? Wie stark ist eine Position wirklich, die auf diese Weise versuchen muss, den freien Zugang zur Gegenargumentation zu blockieren oder zumindest zu erschweren? 51 Der intelligente und vorurteilsfreie Leser wird sich durch solche Methoden nicht davon abhalten lassen, aufgrund der unterbreiteten Tatsachen und Argumente selbst zu einem qualifizierten Urteil zu gelangen. Bei den hoffentlich wenigen Lesern jedoch, denen es statt um Fakten, die Wahrheit und einen rational-ehrlichen, fairen und offenen Diskurs der zahlreichen naturwissenschaftlichen Probleme der Synthetischen (und weiterer) Evolutionstheorie(n), statt der Argumente für und gegen einen intelligenten Ursprung der Lebensformen (intelligentes Design, ID) vielmehr im Sinne des Zeitgeistes um die Frage geht, "wer im Kräftefeld der institutionellen Machtverhältnisse über die größere Definitionsgewalt verfügt und damit auch die Deutungen der Realität mitbestimmen kann" (R. Schmidt 2006, p. 170, vgl. http://www.weloennig.de/RobertSchmidtDesigner.html ), bei Lesern (also), denen es um Ansehen, Vorteile, Macht, Geld, Durchsetzung des Materialismus mit möglichst wortgewaltiger Polemik, Stigmatisierung von "Abweichlern" mit unredlichen Unterstellungen (siehe z. B. den Kommentar von Fritz Poppenberg http://www.dreilindenfilm.de/reaktionen/aw_biologen.htm ), bei Lesern, die für Tatsachen und Logik nicht mehr zugänglich sind, da sie mit den heutigen Evolutionstheorien im Prinzip bereits im Besitz der absoluten Wahrheit sind 52 ("Its essential truth is now universally accepted by scientists competent to judge", Hardin; "Evolution is true and the truth can only make us free", Gould), nämlich der absolut sicheren und ausschließlich materialistisch deutbaren "Tatsache Evolution" 53 , habe ich mit ID natürlich nicht die geringste Chance auch nur auf einen Teilerfolg. (Vgl. zu Tendenzen im heutigen Wissenschaftsbetrieb weiter den aufschlussreichen Kommentar des Journalisten und Kulturkritikers Arno Kleinebeckel 2009, p. 30 zitiert in http://www.weloennig.de/Die_Affaere1.pdf , p. 108. Auch die Themen Soziobiologie und Sozialdarwinismus dürften in diesem Zusammenhang nicht uninteressant sein; siehe z. B. die Ausführungen in http://www.weloennig.de/DieZEITanalyse.html .) 14. "Errare humanum est" 54 und "es irrt der Mensch so lang er strebt" (Goethe, Faust) wir alle machen Fehler, auch wenn wir uns noch so viel Mühe geben, sie zu vermeiden. Wenn Fehler jedoch systematisch auftreten, wie Punkt für Punkt bei der gesamten oben ausführlich diskutierten von MN präsentierten Reihe Roridula gorgonias, Byblis gigantea, Heliamphora tatei, Sarracenia pupurea, Utricularia multifida, sowie in diesem Zusammenhang auch noch zu Genlisea (mit ihrem nicht existierenden Saugmechanismus) und dazu in der nächsten Abstammungsversion von Barthlott et al., die der soeben aufgeführten in mehreren Punkten widerspricht, die zweifelhaften Bindeglieder Pinguicula agnatha und P. utricularioides, dann liegt die Frage nach einer ungewöhnlichen Ursache nahe. Im vorliegende Falle scheint mir die Feststellung von K. F. Meis anwendbar, wenn er in http://www.weloennig.de/Die_Affaere1.pdf pp. 36/37 zu den evolutionstheoretischen "Interpretationen, Extrapolationen und Interpolationen", die "mit einem vorgegebenen atheistischen Ziel arbeiten müssen (geistige Größen haben nur materielle Ursachen) und keine erkenntnisoffene Deutung mehr zulassen", feststellt: "Egal wie das Tatsachenmaterial aussieht, das Ziel steht fest. Entsprechend müssen die Fakten "gebogen", "gestreckt" und "gestaucht" werden, um zur Zielvorgabe zu passen." In diesem Sinne darf man wohl auch mit stark ausgeprägter "selektiver Wahrnehmung" bei einigen ihrer Vertreter rechnen, wobei diese dazu tendieren, alles, was zur Zielvorgabe passt, ungeprüft zu übernehmen und entgegenstehende Tatsachen zu ignorieren (wie oben im Detail exemplifiziert). Das hängt jedoch auch weitgehend von der personalen Integrität und dem ausreichenden Kenntnisstand eines Evolutionstheoretikers zum jeweiligen Thema ab. Hohe personale Integrität (ausgeprägte Wahrheitsliebe, Selbsttreue, Unbestechlichkeit, Rechtschaffenheit, Ganzheit, Kohärenz, Langmut, Gründlichkeit, Selbstbeherrschung, Gerechtigkeit, starke Realitätsbezogenheit) 55 ein Evolutionsbiologe mit diesen Eigenschaften wird selbstverständlich viele Fehler vermeiden und nicht nur die Stärken, sondern auch die Schwächen seiner Position und Hypothesen herausarbeiten. Dem Ideal mehr oder weniger nahe kommen nach meinem Verständnis viele Schriften etwa von Schindewolf, Portmann, Gould, Stanley und anderen Evolutionsbiologen trotz mancher weltanschaulicher Vorgaben 56 . 15. Völlig rätselhaft (oder vielleicht auch nicht) ist mir zur Publikation von MNs Buch Evolution im Fadenkreuz des Kreationismus (2009) die Rolle der Evangelischen Kirche, die doch eigentlich die Aufgabe hätte statt den atheistisch motivierten intellektuellen Krieg gegen eine intelligente Schöpfung mit allen Mitteln, einschließlich materiellen, zu unterstützen die möglichst hoch qualifizierte wissenschaftliche und philosophisch-theologische Verteidigung von Gott, Geist, Intelligenz, Design und Weisheit als direkter Quelle des Universums und des Lebens (auch gegen den materialistischen Zeitgeist) zu vermitteln. Der Herausgeber der Buches (MN) bedankt sich u. a. bei Herrn Hansjörg Hemminger von der Evangelischen Zentralstelle für Weltanschauungsfragen (EZW, Berlin), dafür, dass dieser (wie auch A. Beyer) "das gesamte Buch Korrektur las" und ihn "mit vielen hilfreichen Anmerkungen unterstützte" (d. h. beide haben die zahlreichen Sachfehler des Buches völlig übersehen, für das Utricularia-Kapitel scheinbar ebenso Barthlott) und bei Hemminger auch dafür, dass er ihn "in Verlagsangelegenheiten tatkräftig unterstützte". Darüber hinaus bedankt sich MN für die finanzielle Zuwendung beim evangelischen Oberkirchenrat Stuttgart. ("Für die finanzielle Zuwendung zur Förderung dieses Bandes danke ich meinen Mitautoren sowie dem evangelischen Oberkirchenrat Stuttgart."). Dieser Oberkirchenrat besteht aus der Oberkirchenrätin Margit Rupp, dem Oberkirchenrat Prof. Dr. Ulrich Heckel, Oberkirchenrat Werner Baur, Oberkirchenrat Wolfgang Traub, Oberkirchenrat Erwin Hartmann, Oberkirchenrat Dr. Martin Kastrup und Oberkirchenrat Hans-Peter Duncker 57 . Selbst der schon oben zitierte Hubert Rehm meint zum Druckkostenzuschuss u. a. (2009, p. 55): "Seltsam mutet den Rezensenten auch die Tatsache an, dass das Buchprojekt finanziell vom evangelischen Oberkirchenrat Stuttgart unterstützt wurde. Seit wann sind naturwissenschaftliche Fragestellungen ein Anliegen von Oberkirchenräten?" Der kritische Leser sehe sich jedoch zur offiziellen Position der Evangelischen Kirche einmal die letztlich gegen jede Form einer direkten intelligenter Schöpfung gerichteten, polemischen Schriften der Evangelischen Zentralstelle für Weltanschauungsfragen (Berlin) unter der Federführung von Hansjörg Hemminger an. Wenn nach Hemminger (sinngemäß) von Gott in der Natur überhaupt nichts erkennbar ist (Hemminger 2010, Junker 2010), so steht er damit im diametralen Gegensatz zu Paulus: ". . .weil das, was man von Gott erkennen kann, unter ihnen offenbar ist, denn Gott hat es ihnen offenbar gemacht. Denn seine unsichtbaren [Eigenschaften] werden seit Erschaffung der Welt deutlich gesehen, da sie durch die gemachten Dinge wahrgenommen werden, ja seine ewigwährende Macht und Göttlichkeit, so daß sie unentschuldbar sind;" (Römer 1:19-20) Vgl. weiter zu Hemminger die gut dokumentierten Stellungnahmen von Reinhard Junker und weiterer Autoren: Eine Karikatur des Design-Arguments 58 ; "Mit der Evolution gegen den Bibelfundamentalismus. Stellungnahme der Studiengemeinschaft Wort und Wissen zu: Hansjörg Hemminger: "Mit der Bibel gegen die Evolution Kreationismus und 'intelligent Design' kritisch betrachtet" EZW-Texte 195, 2007 59 ; Und Gott schuf Darwins Welt, Rezension von Reinhard Junker und Henrik Ullrich 60 (2009); dort auch die Links auf die Beiträge von Hemminger. Der evangelische Theologe Dr. theol. habil. Bernhard Kaiser, Leiter des Instituts für Reformatorische Theologie, kommentiert die Position der Evangelischen Kirche zum Thema Schöpfung in seinem Beitrag Die EKD und die Abschaffung der Schöpfung (2008) wie folgt (Hervorhebungen im Schriftbild wieder von mir): "Es ist ein Skandal, daß die neuerlichen kirchlichen Publikationen die jahrzehntelange Tradition fortsetzen, anstelle der biblischen Botschaft widerbiblische und dem jeweiligen Zeitgeist angepaßte Positionen vorzutragen. Die Kirche sollte doch, um in rechter Weise Kirche zu sein, dem Wort Gottes mehr glauben als einer mit dem Schein der gesicherten Erkenntnis sich schmückenden naturalistisch überhöhten Wissenschaft. Wenn sie darüber hinaus die recht verstandene biblische Schöpfungsaussage diffamiert als "Verkehrung des Glaubens an den Schöpfer in eine Form einer Welterklärung, die letztlich dazu führt, daß das Bündnis von Glaube und Vernunft aufgekündigt wird" (Beintker/Friedrich 7), dann tut sie genau das, was sie kritisiert: sie hat mit der Rede von den verschiedenen Ebenen die biblische Schöpfungsaussage als eine vernünftige und sachlich richtige Auskunft über die Herkunft der Welt aus der Hand eines weisen Schöpfers von der Wissenschaft abgekoppelt" vgl. http://www.wort-und-wissen.de/index2.php?artikel=disk/d08/3/d08-3.html . Siehe zur Anpassung an den jeweiligen Zeitgeist das folgende (Muster-) Beispiel aus der neueren deutschen Geschichte, das Verhalten der Evangelischen Kirche während der NS-Zeit: http://de.wikipedia.org/wiki/Stuttgarter_Schuldbekenntnis . Kurzzitate: "Durch uns ist unendliches Leid über viele Länder und Völker gebracht worden." Oder aus Predigt von Hans Christian Asmussen: "Schuldig ist die Kirche beider Konfessionen. Wir haben durch lange Jahrzehnte versucht, mit Weltanschauungen zu paktieren, für welche es keine letzte Wahrheit gibt. Anstatt zu sagen ,Nein', haben wir gesagt ,Sowohl-als-auch." Weiteres zum Thema Religion im Beitrag http://www.weloennig.de/Die_Affaere1.pdf .
Literatur (nur Ergänzungen die übrige Literatur siehe unter Die in den Links direkt anklickbaren Artikel werden hier meist nicht noch einmal aufgeführt
Anderson, B. (2005): Adaptations to foliar absorption of faeces: a pathway in plant carnivory. Annals of Botany 95: 757-761. Benzinq, D. H. (1987): The origin and rarity of botanical carnivory. Trends in Ecology and Evolution 12: 364-369. Darnowski, D. W., Carroll, D. M., Plachno, B. and E. Cinnamon (2006): Evidence of Protocarnivory in triggerplants (Stylidium spp.; Stylidiaceae). Plant Biology 8: 805-812. Evert, R. F. (2006): Esaus's Plant Anatomy. Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development. Third Edition. John Wiley & Sons. Frahm, J. P. und J. Eggers (2001): Lexikon deutschsprachiger Bryologen. Books on Demand, Norderstedt. Hartmeyer, I. und S. Hartmeyer (2005) Das Taublatt: http://www.hartmeyer.de/ArtikelundBerichte/artBfiliEnzym.htm Heslop-Harrison Y. und J. Heslop-Harrison (1980): Chloride ion movement and enzyme secretion from the digestive glands of Pinguicula. Annals of Botany 45: 729-731. Heslop-Harrison, Y und J. Heslop-Herrison (1981): The Digestive Glands of Pinguicula: Structure and Cytochemistry. Annals of Botany 47: 293-319. Junker, T. (1989): Darwinismus und Botanik. Rezeption, Kritik und theoretische Alternativen im Deutschland des 19. Jahrhunderts. Mit einem Geleitwort von Rudolf Schmitz. In Kommission: Deutscher Apotheker Verlag, Stuttgart. Lambers, H., Chapin, F. S. and T. L. Pons (2008): Plant Physiological Ecology. 2nd edition. Springer Verlag, Berlin. Lang, F. X. (1901): Untersuchungen über Morphologie, Anatomie und Samenentwicklung von Polypompholyx und Byblis. Flora 88: 149-206. (Die Arbeit enthält genaue Beschreibungen, aber bedauerlicherweise auch einige grobe Fehler nicht weil die Arbeit so alt ist, sondern weil der Verfasser im Sinne des Darwinismus Ableitungen versucht, die nicht zutreffen.) Lloyd, F. E. (1936): Notes on Utricularia. With special reference to Australia, with descriptions of four new species. The Victorian Naturalist 53: 91-112. P. 104: "The actual entrance is small. The lower part of the door lies curved downwards over a narrow ridge of the threshold in the set posture. After actuation the door is also curved downward in front of the threshold. There is a particular velum. The effective blocking of in-leakage being brought about by the supplementary action of numerous long glandular hairs. In transverse section the trap is seen to be three-sided, quite different in this from all other utricularias. When the trap is set, the walls are concave, more especially the side walls, the top being less responsive." Wie oben schon nach Juniper et al. und anderen zitiert, war Lloyd ein Meister der genauen Beobachtung und anatomischen Beschreibungen ("No purpose would be served by repeating all his [Lloyd's] observations or his conclusions, few if any of which need qualification even after the passage of 50 years."). Von Roridula kannte er jedoch nur Material in Formalin und kommentiert (1942, p. 5, ähnlich p. 98): "The leaves bear many tentacles superficially similar to those of Drosera. Examination showed them to be anatomically quite different, and that they exude a resinous secretion. There are no other glands [soweit alles korrekt], so that on this evidence the carnivorous habit seems to be quite excluded (LLOYD, 1934)." Von den oben referierten extrazellulären Phosphatasen und den Untersuchungen Andersons zur speziellen Struktur der Epidermis sowie der massiven "mineral uptake (N, P, K and Mg) von Drosophila prey bei Roridula auch ohne Symbionten" konnte er natürlich noch nichts wissen. Mayr, E. (2001): What Evolution is. Basic Books, New York. Zitiert nach http://www.evolutionnews.org/2009/05/a_primer_on_the_tree_of_life_p_2020161.html Müller K., Borsch, T., Legendre, L., Theisen, I. und W. Barthlott (2006): Evolution of carnivory in the Lentibulariaceae: considerations based on molecular, morphological, and physiological evidence. Presentation at the International Carnivorous Plant Society meeting in Tokyo (2002). Neukamm, M. (18. 5. 2010): Eine unendliche Geschichte: Dr. W.-E. Lönnig, Intelligent Design und die Saugfalle der Pflanze Utricularia vulgaris. [Untertitel:] Die Evolution der karnivoren Pflanzen: Was die Selektion in vielen Einzelschritten zu leisten vermag. Oder: Weshalb Dr. Lönnig den Vaterschaftstest nicht verstanden hat: http://www.martin-neukamm.de/loennig-utricularia.pdf (letzter Zugriff am 20. 3. 2011) Kurzer Ergänzungskommentar: Ich könnte nun wie oben für seinen Beitrag "Was die Selektion angeblich nicht leisten kann. Diskussion von drei Paradebeispielen." "1: Die Saugfalle des Wasserschlauchs Utricularia" (2009, pp. 240-250) zu 112 Einzelpunkten durchgeführt auch seinen weiteren polemischen Beitrag von 2010 in allen Details analysieren (das ergäbe schätzungsweise 400 Seiten Text). Der vorurteilsfreie Leser wird jedoch schon anhand der wenigen oben aufgeführten Punkte zur " unendlichen Geschichte " in der Lage sein, sich ein Bild von den Methoden und der Wissenschaft auch dieses Beitrags von MN zu machen, einmal ganz abgesehen von den ad hominem-Attacken, die man wohl zum großen Teil wieder unter dem Begriff unbeabsichtigte aber sehr treffende Selbstbeschreibung subsumieren kann, vgl. p. 4 von http://www.weloennig.de/Die_Affaere1.pdf , einschließlich der von weiteren Personen gebrauchten Begriffe wie "Philippika" ("bezeichnet eine Angriffs-, Brand-, oder Kampfrede") sowie die mir unterstellte "Unbelehrbarkeit". Stil und Methode mancher meiner Kritiker erinnert vielleicht an die Art und Weise, mit der K. F. Schimper ("ein ausgesprochen vielseitiger Gelehrter" T. Junker) einst den Darwinismus zurückgewiesen hat: "Die Zuchtlehre Darwins ist, wie ich gleich gefunden, und bei wiederholtem aufmerksamem Lesen nur immer besser wahrnehmen mußte, die kurzsichtigste, niedrig dummste und brutalste die möglich und noch weit armseliger als die von den zusammengewürfelten Atomen, mit der ein moderner Possenreißer und gemieteter Fälscher bei uns sich interessant zu machen versucht hat." (Zitiert nach T. Junker 1989, p. 41 und Frahm/Eggers Lexikon deutschsprachiger Bryologen 2001, p. 452. T. Junker kommentiert dieses Zitat u. a. mit der Fußnote 142 (ebenfalls p. 41).: "Mit dem 'modernen Possenreißer' und gemieteten Fälscher ist wohl der Materialist Karl Vogt gemeint, der ein bezahlter Agent Napoleons III gewesen sein soll. Vgl. Wittich I (1971), XXI bzw. Lauterborn (1934), 304 Fn.") Interessanterweise scheinen sich einige Vertreter der Evolutionstheorie bis auf den heutigen Tag noch über Schimpers Polemik von 1865 aufzuregen und übersehen dabei, dass sie mit zweierlei Maß messen, wenn sie selbst ähnlich verfahren bzw. Schimpers Methode durch persönliche Angriffe und mangelndes Studium der Intelligent-Design-Theorie (ID) sogar noch übertreffen. Denn zahlreiche Unterstellungen und unzutreffende Behauptungen zur ID-Theorie sowie zu meinen Arbeiten sind eigentlich nur möglich, wenn man diese nicht gründlich studiert hat 61 . Schimper hatte sich hingegen der "Zuchtlehre Darwins" wenigstens noch mit "wiederholtem aufmerksamem Lesen" gewidmet und wusste daher genau, wovon er sprach. Zu den mehr als fragwürdigen Methoden vieler Kritiker im Umgang mit Befürwortern des intelligenten Ursprungs der Lebensformen, möchte ich die daran interessierten Leser noch einmal ermuntern, sich sehr genau das Dokument "DIE AFFÄRE MAX PLANCK", DIE ES NIE GEGEBEN HAT http://www.weloennig.de/Die_Affaere1.pdf anzusehen. MN hat übrigens seinen oben behandelten Buchbeitrag "Was die Selektion angeblich nicht leisten kann. Diskussion von drei Paradebeispielen." Unterkapitel "1: Die Saugfalle des Wasserschlauchs Utricularia" (2009), den man unter dem Link http://www.evolution-im-fadenkreuz.info/KapIX_1.pdf unmittelbar überprüfen konnte, aus dem Internet genommen. Stattdessen fand man eine Zeitlang den Beitrag von Johannes Sikorski "Die bakterielle Flagelle: Aufbau, Diversität und Evolution", der inzwischen ebenfalls aus dem Netz genommen wurde. Es kann daher gut sein, dass MN auch sein fehlerreiches Dokument von der "unendlichen Geschichte" in absehbarer Zeit wieder aus dem Netz nimmt. Mit scheint es zurzeit sinnvoller, die Diskussion hier zu unterbrechen und selbst weiter in die empirische Forschung zu gehen und zu einigen Punkten eigene Untersuchungen zu den Karnivoren am lebenden Pflanzenmaterial durchzuführen. Zum Thema Vaterschaftstest siehe zum Beispiel: Abstammungsgutachten: http://de.wikipedia.org/wiki/Abstammungsgutachten_(DNA-Analyse) und http://de.wikipedia.org/wiki/Abstammungsgutachten . Pohl, S. (2009): Untersuchungen zur möglichen Protokarnivorie von Lathraea squamaria, Salvia glutinosa und Rubus phoenicolasius. Diplomarbeit, Universität Wien. Fakultät für Lebenswissenschaften. Abstract: Die drei in Österreich vorkommenden Pflanzenarten Lathraea squamaria, Rubus phoenicolasius und Salvia glutinosa weisen Strukturen auf, die eine Zuordnung zu den protokarnivoren Pflanzen vermuten lassen. L. squamaria hat dicht mit Drüsen besetzte Aushöhlungen in ihren unterirdischen Niederblättern, die an die Fallen von Genlisea erinnern. Bei R. phoenicolasius und S. glutinosa sind gewisse Pflanzenteile mit klebrigen Drüsen besetzt, ähnlich den Klebfallen karnivorer Pflanzen (z.B. Drosera). Zudem leben auf S. glutinosa verschiedene Wanzen, mit denen eine ähnliche Symbiose bestehen könnte wie bei der Protokarnivoren Roridula. In dieser Arbeit wurde durch Untersuchungen am Naturstandort, kontrollierte Fangversuche, makro- und mikroskopische Betrachtung der Fangblätter und ihrer Drüsen sowie durch cytochemische Nachweise versucht, Hinweise für oder gegen eine mögliche Protokarnivorie dieser Pflanzen zu finden. Die Menge der im Boden verfügbaren Nährstoffe zeigt keine Notwendigkeit einer zusätzlichen Ernährung durch eventuelle Beutetiere. Ein Fangvermögen der möglichen Fallen ist am jeweiligen Naturstandort auf jeden Fall gegeben, unter Laborbedingungen ist die Wahl der angebotenen Beute entscheidend für das Ergebnis. Die Schuppenblätter von L. squamaria speichern massenhaft Stärke, jedoch keine Proteine. Die Drüsen in den Aushöhlungen der Blätter produzieren keine Verdauungsenzyme und sind auch nicht geeignet, um gelöste Stoffe aufzunehmen. Ihre Aufgabe ist es, den Wasserhaushalt der Pflanze zu kontrollieren. In R. phoenicolasius gibt es keine Speichergewebe. In den Drüsenstielen sind Gerbstoffe eingelagert, die als Fraßschutz dienen. Die vielzelligen Drüsenköpfchen geben einen wasserlöslichen Schleim ab, der keine Verdauungsenzyme enthält. Eine Aufnahme wasserlöslicher Nährstoffe konnte nicht festgestellt werden. Die klebrigen Pflanzenorgane dienen zum Schutz der Knospe. Auch in S. glutinosa treten keine Speichergewebe für Stärke oder Proteine auf. Die Köpfchen der gestielten Drüsen am Blütenstand sind einzellig und sezernieren ein lipidhaltiges, harziges Sekret. Es enthält keine Verdauungsenzyme und dient vermutlich hauptsächlich der Anlockung von Bestäubern. Wenn eine Symbiose von S. glutinosa mit den auf ihr gefundenen Wanzen besteht, so ist noch nicht geklärt, wie die Nährstoffe in die Pflanze gelangen. Über die Blütenstände oder die Laubblätter können keine wässrigen Flüssigkeiten aufgenommen werden. Aufgrund der Ergebnisse dieser Arbeit ist keine der drei Pflanzen als protokarnivor zu bezeichnen, da sie offenbar keinen Nutzen von den gefangenen Tieren haben. Siehe: http://othes.univie.ac.at/6995/ Rokas, A, Krueger, D. and S. B. Carroll (2005): Animal evolution and the molecular signature of radiations compressed in time. Science 310: 1933-1938. Slack, A. (2006): Insect eaters. How to grow and feed extraordinary plants from the Venus Fly Trap to the Cobra Lily. Alphabet and Image Publishers, Totnes, Devon. Printed and bound in China by Compass Press. (P. 102: Genlisea is a strange semi-aquatic plant which resembles no other carnivorous genus in trap structure, although it is closely related to the bladderworts Utricularia.) Spomer, G. G. (1999): Evidence of protocarnivorous capabilities in Geranium viscosissimum and Potentilla arguta and other sticky plants. International Journal of Plant Sciences 160: 98-101. Vassilyev , A. E. und L. E. Muravnik (1988): The ultrastructure of the digestive glands in Pinguicula vulgaris L. (Lentibulariaceae) relative to their function. II. The changes on stimulation. Annals of Botany 62: 343-351. Watchtower Bible and Tract Society (1981): Accidents of Evolution or Acts of Creation? Awake! (22. 9. 1981; deutsche Ausgabe 22. 12. 1981, Kasten p. 21: DIE "TYRANNEI DER AUTORITÄT" VON SEITEN DER EVOLUTIONISTEN, "Als er [Darwin] abgeschlossen hatte, konnte die Tatsache der Evolution nur durch ein Aufgeben der Vernunft geleugnet werden" (Life Nature Library, "Evolution", S. 10). "Es ist keine Frage des persönlichen Geschmacks, ob wir an die Evolution glauben oder nicht. Die Beweise für die Evolution sind zwingend" ("Evolution, Genetics, and Man", S. 319, Dobzhansky). "Ihre grundlegende Wahrheit wird heute generell von Wissenschaftlern akzeptiert, die kompetent sind, sich ein Urteil zu bilden" ("Nature and Man's Fate", p. v, Hardin). "Der Aufbau vom Familienbaum des Lebens durch den Prozeß der Evolution wird heute von allen verantwortungsbewußten Wissenschaftlern anerkannt" ("A Guide to Earth History", S. 82, Carrington). "Niemand, der informiert ist, leugnet heute, daß der Mensch durch einen langsamen Prozeß aus der Welt des Fisches und des Frosches entstanden ist" (Zeitschrift "Life", 26. August 1966, Ardrey). "Es ist heute schon fast selbstverständlich geworden und erfordert keine weiteren Beweise für jemand, der in einem vernünftigen Rahmen frei ist von alten Illusionen und Vorurteilen" ("The Meaning of Evolution", S. 338, Simpson). "Es gibt keine konkurrierende Hypothese außer der veralteten und völlig widerlegten Lehre von der direkten Erschaffung, die heute nur noch von unwissenden, dogmatischen und voreingenommenen Personen aufrechterhalten wird" ("Outlines of General Zoology", S. 407, Newman).") Woese, K. (1998): The universal ancestor. PNAS 95: 6854-6859. "Phylogentic incongruities can be seen everywhere in the universal tree, from its root to the major branchings within and among the various taxa to the makeup of the primary groupings themselves. |
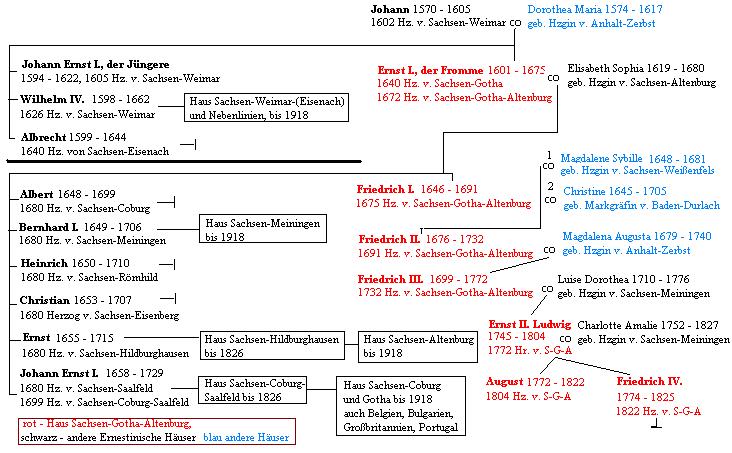
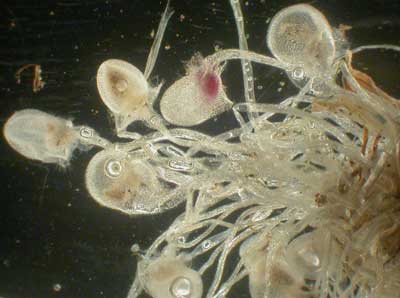



1 Box THE "TYRANNY OF AUTHORITY USED BY EVOLUTIONISTS, Awake! 22 September 1981, p. 21; ähnliche Zitate in den Ausgaben vom Januar 1990 und September 2006; siehe weiter den Beitrag von Robert Schmidt (2006): "Götter und Designer bleiben draußen ": http://www.weloennig.de/RobertSchmidtDesigner.html , Lönnig 2001/2007: http://www.weloennig.de/Utricularia.html und 2010 http://www.weloennig.de/Die_Affaere1.pdf ,, sowie die Einleitungen zu meinen naturwissenschaftlichen Vorträgen unter http://www.unser-auge.de/intelligent-design-hypothese/intelligent-design_ursprungsfrage_der_biologie.html , insbesondere vielleicht http://www.unser-auge.de/intelligent-design-hypothese/intelligent-design_kassel_1.html . 2 Ja, die Probleme sind sogar so tiefgreifend, dass einige Personen behaupten, die Klärung der Einzelschritte sei ja keinesfalls das Anliegen des von mir oben im Detail analysierten Unterkapitels zu Utricularia. 3 Siehe z. B. http://www.weloennig.de/Antwort_an_Kritiker.html 5 "Für ihn ist die Darwinsche Evolutionstheorie eine Ersatzreligion, die es gegen jegliche (also auch sachliche) Kritik zu verteidigen gilt" - Georg Menting, vgl. weiter die Fußnote 205, p. 117, insbesondere auch die Ausführungen zu MNs Umgangsmethoden. 6 Formulierung in Anlehnung eines Beitrags von Casey Luskin (2009): http://www.evolutionnews.org/2009/05/does_any_critic_out_there_unde020491.html . Die Ambivalenz eines solchen Unterfangens wurde jedoch schon vor etwa 3000 Jahren von König Salomo im Buch der Sprüche, Kapitel 26, Verse 4 und 5 beschrieben (siehe dazu eine moderne Übersetzung wie die NW: "Antworte nicht /Antworte damit"). 7 Vgl. http://www.weloennig.de/Die_Affaere1.pdf (insbesondere pp. 79 und 100). 8 - wie ihn seine Kollegen von der Redaktion beschreiben; siehe Laborjournal 7-8/2010, p. 3. 9 - die in weniger als 1/1000stel Sekunde 'zuschnappt'; Arzt 2008 und Vincent et al. 2011; nach Slack 2006, p. 104 "in something in the region of a ten-thousandth to fifteen-thousandth of a second". 10 Vgl. zum Flagellenapparat die differenzierten Ausführungen unter http://www.evolutionnews.org/2011/03/michael_behe_hasnt_been_refute044801.html und Scherer 2010 http://www.evolutionslehrbuch.info/teil-4/kapitel-09-04-r01.pdf 11 Vgl. dagegen die Arbeiten von Lönnig: http://www.weloennig.de/AuIn.html , Ullrich, Winkler und Junker http://www.si-journal.de/index2.php?artikel=jg13/heft1/sij131-1.html und Ullrich http://www.wort-und-wissen.de/index2.php?artikel=sij/sij151/sij151-4.html sowie Berlinski http://www.discovery.org/a/1416 und dazu die ausführliche Diskussion unter http://www.discovery.org/a/1509 12 Vgl. Laborjourmal 12/2009, pp. 54-55: "Die Reduktion des Irreduziblen"; siehe auch das Zitat oben pp. 108/109. 13 - von den mehr als zweifelhaften ad-hominem-[gegen eine Person gerichteten]Attacken einmal ganz abgesehen. 14 http://www.laborjournal.de/rubric/archiv/domfac/bellbio/schoen_10_02.pdf 15 Zur Rechtfertigung der widersprüchlichen Stammbäume zur Makroevolution gibt es dann auch jede Menge z. T. auch vom Evolutionstheoretiker R. Neumann aufgeführte nicht oder kaum nachprüfbarer evolutionärer Deutungen: evolutionäre Tempounterschiede für spezifischen Anpassungen, Konvergenzerscheinungen, einige Gene eignen sich schlecht zur Konstruktion phylogenetischer Bäume etc. Können solche Überlegungen, die in der Regel die Gesamtevolution wieder als nicht hinterfragbare Theorie voraussetzen, tatsächlich die vielen erstaunlichen Widersprüche erklären? Gibt es vielleicht eine bessere Erklärung für diese Phänomene? 16 Ein paar Punkte davon wurden schon auf der Seite 125 oben erwähnt; Schriftbild von mir verändert. 17 http://www.evolutionnews.org/2011/04/who_misrepresented_who_a_respo045471.html (Schriftbild zumeist von mir.) 18 Selbst die verschiedenen Exons von ein und demselben Gen können zu widersprüchlichen Stammbäumen führen; Anm. von W-EL (so geschehen in einer Dissertation, die ich hier nicht näher nennen möchte). 19 Ganz seltene Ausnahme (um die 30 Fälle unter zig Millionen): Chimären, die jedoch letztlich ebenfalls eindeutig genetisch verstanden werden können (vgl. http://abcnews.go.com/Primetime/story?id=2315693 "In human biology, a chimera is an organism with at least two genetically distinct types of cells -- or, in other words, someone meant to be a twin. But while in the mother's womb, two fertilized eggs fuse, becoming one fetus that carries two distinct genetic codes -- two separate strands of DNA. 20 In der forensischen Medizin wird selbstverständlich kein Zirkelschluss aufgrund von Ähnlichkeiten praktiziert, sondern hier liegt eine über 99 prozentige Sicherheit aufgrund des genetischen Fingerabdrucks zur Feststellung der Identität (nicht der Ähnlichkeit) vor (Fehlerquote in den Jahren 1998-2002 bei 0,4 0, 7 %; siehe http://www.biospektrum.de/blatt/d_bs_download&_id=973596 ). "Allerdings ist ein alleiniger "DNA-Beweis" unter juristischen Aspekten für eine Verurteilung nicht ausreichend." "Wie jedes Verfahren hat auch die forensische DNA-Analyse Grenzen" - E. Reuss 2008, pp. 709 und 710 ( http://www.biospektrum.de/blatt/d_bs_download&_id=973596 ). Weitere Erhöhung der Sicherheit durch ergänzende Verfahren zur Identifikation eines Täters mit in der Regel anschließenden Schuldbekenntnissen, oft begleitet von Zeugenaussagen. 21 Die endlos repetierte Aussage, die hypothetisch-deduktive Methode und damit die Naturwissenschaft überhaupt könne grundsätzlich nur zu vorläufigen Ergebnissen führen (womit der Einwand vom Zirkelschluss gegenstandslos sei), steht dabei im schärfsten Gegensatz zur stereotyp-suggestiv wiederholten Behauptung, die (Makro-)Evolution sei eine absolut sichere Tatsache, die kein vernünftiger Mensch mehr bezweifeln könne etc. (vgl. oben die einleitende Zitate zur hier vorliegenden Diskussion von Einwänden). Es geht also beim Thema Evolution nicht nur um hypothetische Erklärungen ohne Anspruch auf Realität, sondern um die Realität und die Tatsachen selbst. Auf dieser Ebene fehlen jedoch die Beweise und hier gilt der oben diskutierte Zirkelschlusseinwand. 22 Siehe im Folgenden; es gibt jedoch weitere Ansatzpunkte und Deutungsmöglichkeiten und scheinbar mehrere strittige Punkte in dieser Beschreibung der hypothetisch-deduktiven Methode, auf die ich hier jedoch nicht weiter eingehen möchte. 23 Laborjournal 5/2009 http://www.laborjournal.de/rubric/archiv/domfac/bellbio/schoen_09_05.pdf 24 http://www.evolutionnews.org/2011/03/a_closer_look_at_one_scientist045311.html 25 http://www.weloennig.de/Gesetz_Rekurrente_Variation.html 26 - und der auch nach der Theorie grundsätzlich unbeobachtbar ist; vgl. Dokumentation unter http://www.weloennig.de/Gesetz_Rekurrente_Variation.html speziell die Zitate von Schindewolf, Dobzhansky, Andermann. 27 Bedeutender deutscher Botaniker (Hauptarbeitsgebiet Paläobotanik), der "einen abrupten Wechsel der einzelnen Floren in der Tradition von Cuvier zu vertreten scheint. Göppert lehnt sowohl einen genetischen Zusammenhang der Arten als auch die Evolution ab. Den Ursprung der Arten vermutet er vielmehr in getrennten Schöpfungsakten, wobei die verschiedenen Formen durch einen 'schöpferischen Typus' bewirkt sein sollen" T. Junker 1989, p. 97. 28 Juniper, Robins und Joel z. B. bemerken in ihrem Meisterwerk The Carnivorous Plants zu Lloyds Utricularia-Studien 1989, p. 66 u. a.: "No purpose would be served by repeating all his observations or his conclusions, few if any of which need qualification even after the passage of 50 years." 29 Wertlosigkeit aller exakten naturwissenschaftlichen Beschreibungen, die sagen wir jeweils älter als 50 Jahre sind. 30 On Polypompholyx (2001, p. 138): "Like Utricularia spp., the prey is sucked in when the trigger hairs are stimulated. 31 http://www.waltraudschulze.de/carnivor/suction.html 32 http://www.life.umd.edu/grad/mlfsc/Carnivorous%20Plants.pdf 33 http://www.britannica.com/EBchecked/topic/229188/Genlisea 34 http://www.inet1000.com/IPW_BOOK/Text.htm 35 Darwin (1875) spricht hingegen wiederholt von einer "slitlike orifice bei verschiedenen Utricularia-Arten (Polypompholyx hat er jedoch nicht in seine Studien mit einbezogen). Er zeichnet die Tür jedoch anliegend (Fig. 20) und sagt unter anderem (p. 406): "Animals enter the bladders by bending inwards the posterior free edge of the valve, which from being highly elastic shuts again instantly. As the edge is extremely thin, and fits closely against the edge of the collar, both projecting into the bladder (see section, fig. 20), it would evidently be very difficult for any animal to get out when once imprisoned, and apparently they never do escape. To show how closely the edge fits, " 36 aff.: "The term affinis (or sp. aff = species affinis) is used when the identity of a distinct biological species is unknown but it has a striking similarity or close relation with a known species " vgl. http://www.cactus-art.biz/note-book/Dictionary/Dictionary_S/dictionary_species_affinis.htm ; Zugriff auf die letzten 4 Links am 16. März 2011. 37 Vgl. zu den funktionalen und selektionstheoretischen Schwierigkeiten der Idee submerser Utricularia-Fallen ohne Saugmechanismus auch die Ausführungen zu den Punkten 38, 39 und 42, pp. 51-53 der vorliegenden Arbeit. 38 Allen Lowrie vermutet hingegen "Evolution at work mit den unwahrscheinlichen Vielfach-Konvergenzerscheinungen (vgl. pp. 73, 124) ist dieser Ansatz jedoch nicht unproblematisch. Forschungsaufgabe: Mayrs Begriff "hidden potentials of the genotype" molekular fassen. 39 " two American species of sticky plants suspected of carnivory, namely Ibicella lutea and Proboscidea parviflora (Martyniaceae), and in two semi-desert species with glandular sticky leaves, Cleome droserifolia (Capparidaceae) and Hyoscyamus desertorum (Solanaceae), sowie Drosophyllum lusitanicum (Droseraceae) und Roridula gorgonias (Roridulaceae) 40 http://en.wikipedia.org/wiki/Protocarnivorous_plant . 41 Vgl. auch http://en.wikipedia.org/wiki/Protocarnivorous_plant 42 - "...virtually every surface of a plant is capable of absorbing both large and small molecules" Juniper 1986 zitiert nach Spomer 1999, p. 98. Das ist jedoch im allgemeinen umgekehrt proportional zur Molekülgröße, d. h. je größer die Moleküle desto schwieriger und geringer die Aufnahme; Analogie: auch die menschliche Haut ist zur selektiven Absorption von Molekülen befähigt (Stichwort "Nikotinpflaster") ist damit die Entstehung des synorganisierten Multikomponentensystems des Verdauungssystems mit Mund, Zunge, Speicheldrüsen, Zähnen, Magen, Leber, Gallenblase, Pankreas bis zum Anus durch definitionsgemäß richtungslose Mutationen und Selektion erklärt? Kann das wunderbare Organ Haut mit den ihr eigenen komplexen Strukturen und vielfältigen Aufgaben die specified complexity des Verdauungskanals ersetzen oder auch nur ausreichend funktional erklären? Aber zurück zu Aufnahme kleinerer Moleküle über das Blatt: "Die Aufnahme von gelösten Stoffen über die Blätter ist bei Landpflanzen durch die Cuticula der Epidermis stark eingeschränkt. Niedermolekulare Verbindungen wie Zucker sowie Mineralstoffe und Wasser können durch hydrophile Poren die Cuticula passieren. Diese Poren haben einen Durchmesser von einem Nanometer , dadurch kann z. B. Harnstoff (Durchmesser 0,44 Nanometer) leicht passieren. Die Poren sind negativ geladen, so dass Kationen leichter passieren können als Anionen . Damit wird z. B. Ammonium rascher aufgenommen als Nitrat " http://de.wikipedia.org/wiki/Blatt_(Pflanze) . " Zu den Nachteilen der Blattdüngung zählen zum Beispiel [wieder nach soeben zitierten Quelle]: 1. Abperlen von der hydrophoben Blattoberfläche 2. Abwaschen durch Regen 3. Bestimmte Nährstoffe wie Kalzium können von den Blättern nicht mehr in andere Pflanzenteile transportiert werden. 4. Mit einer Blattdüngung kann nur eine begrenzte Menge an Nährstoffen
aufgebracht werden (Ausnahme ist Harnstoff). Wollte man jedoch sämtliche Pflanzenarten, die in der Lage sind, zumindest kleinere Moleküle über das Blatt aufzunehmen, als para- oder protokarnivor einstufen, dann müsste man praktisch das gesamte Pflanzenreich zu den Para- oder Protokarnivoren zählen. Aufgenommen werden am besten relativ kleine Moleküle wie N über Harnstoff (CH4N2O), Phosphor in H3PO4, Kalium in KNO3 usw. (vgl. Tabelle 3 in http://www.conklin.com/files/pdf/ag0100_006_0909_mwlabsfoliarnutrition.pdf ). Siehe dagegen die Bedingungen für das CARNIVOROUS SYNDROME unten, wobei jeder einzelne Schritt zum Syndrom ein unwahrscheinliches Ereignis für sich ist. 43 Der Punkt 2 in der Auflistung von Honda wird von anderen Autoren in zwei Extrapunkte unterteilt: "2. Capture of prey, "3. Retention of prey und vor "Digestion of prey" kommt "Killing auf prey. Zum Punkt 4 nach Honda wäre "in substantial amounts zu ergänzen und vor "Derivation of benefit" könnte man "Translocating the nutritive yield auführen (Benzinq nach Lloyd und Heslop-Harrison) und zu "Derivation of benefit selbst könnte man "i. e. clear use for growth and reproduction" ergänzen. Mit Nachtwey hatten wir oben noch den Schritt Umwandlung der tierischen Proteine in "arteigenes Planzeneiweiß" zitiert. Hier dürfte zur generellen Fähigkeit aller Pflanzen zur Kodierung und Produktion von Proteinen bei den Karnivoren zwecks Abbau und Umbau tierischer Proteine noch mindestens ein Schritt dazu kommen (die oben erwähnten Spezies der Gattungen Proboscidea, Ibicella, Cleome und Hyoscyamus können das offenbar nicht; Forschungsaufgabe: genau welche(r) Schritt(e)?). Das ergäbe dann also 9 Punkte. 45 http://www.evolutionnews.org/2006/04/do_car_engines_run_on_lugnuts002075.html , http://www.evolutionnews.org/2006/04/do_car_engines_run_on_lugnuts_1002157.html, http://www.discovery.org/a/3718 , 46 http://www.weloennig.de/AuIWa.html 47 http://www.weloennig.de/Giraffe_Erwiderung.1a.pdf und http://www.weloennig.de/GiraffaZweiterTeil.pdf ; 48 Einschließlich Ablenkungsmanövern, wie dass bei S. purpurea "Ab dem 2. Jahr die Verdauung allerdings von den Destruenten (z. B. Bakterien) übernommen wird, welche die Falle besiedeln" jetzt schließt er also für das zweite Jahr die (induzierbare) Produktion von Verdauungsenzymen aus anstatt sich zu vergewissern, ob diese Frage überhaupt schon wissenschaftlich untersucht worden ist. Gallie schrieb mir auf meine Anfrage "Is there anything known about induction and enzyme production in the second year, say in one and a half year old pitchers? am 6. 4. 2011: "..we haven't looked at enzyme production in one and a half year old pitchers so we just don't know. My guess is that pitchers remain responsive but perhaps less so as they age. Dafür könnte auch die folgende Beobachtung von Barry Rice sprechen: " the pitchers of Sarracenia purpurea survive in fine shape for two years unless they are damaged by fire or truly excessively cold conditions ( http://www.sarracenia.com/faq/faq5538.html ). MN sagt weiter, dass die Beschreibung in der Karnivoren-Datenbank zur Arbeit von Gallie und Chang nicht eindeutig war anstatt sich die Originalarbeit anzusehen! Und angesichts der oben analysierten fragwürdigen Rechtfertigungsversuche darf man vielleicht auch fragen, ob MN inzwischen die wirklich painstaking investigations von Gallie und Chang (mit ihren zahlreichen Wiederholungen der Einzelversuche zur möglichst einwandfreien Absicherung der Daten und Ergebnisse) gründlich studiert und adäquat verarbeitet hat. 49 Obwohl eine Differenzierung für das erste und zweite Jahr gar nicht zur Diskussion stand und der postulierte Verlust der Fähigkeit im zweiten Jahr für die grundsätzliche Frage, ob S. purpurea überhaupt Verdauungsenzyme produziert, letztlich irrelevant ist. Sollte es sich (entgegen selbst selektionstheoretischer Erwartungen) herausstellen, dass die Verlusthypothese zutrifft, dann könnte die Antwort auch für Degenerations- und Funktionsverlustmöglichkeiten sowohl bei anderen Karnivoren (wie Heliamphora) als auch im Organismenreich generell relevant werden (und damit auch wieder für die mangelnde Stringenz der Selektion überhaupt, siehe oben); vgl. weiter http://www.weloennig.de/AesVI.1Dege.html . 50 Versuchsbeginn: Donnerstag 12. August 2010, Versuchsende Mittwoch 22. September 2010. Den (Pilot-)Versuch müsste man mit verschiedenen Pinguicula-Arten und unterschiedlichen Böden mit größeren Pflanzenzahlen erweitern. 51 Bei meinen Hinweisen auf die materialistische Philosophie mancher Autoren sieht die Situation dagegen völlig anders aus, weil die große Mehrheit des Publikums diese bejaht. 52 Und in der Biologie ist von den Vertretern dieser Auffassungen tatsächlich und eindeutig die naturwissenschaftliche Ebene gemeint. 53 Siehe die Zitate und Links in der Einleitung zur Diskussion von Einwänden oben. 54 - "irren ist menschlich" und es geht noch weiter http://de.wiktionary.org/wiki/errare_humanum_est 55 Z. T. nach P. Büche aus Pollmann, vgl http://de.wikipedia.org/wiki/Integrit%C3%A4t_%28Ethik%29 56 Zu Gould siehe jedoch Fußnote 62, p. 36 oben. 57 Gemäß https://www.service.elk-wue.de/oberkirchenrat/uebersicht-oberkirchenrat.html ; Zugriff am 21. 3. 2011. 58 http://www.wort-und-wissen.de/disk/d10/1/d10-1.pdf 59 http://www.wort-und-wissen.de/disk/d08/2/d08-2.pdf 60 http://www.wort-und-wissen.de/info/rezens/b38.pdf 61 Weitere Möglichkeiten möchte ich an dieser Stelle nicht erwähnen. |
| Zurück zur Homepage |
© Copyright by Wolf-Ekkehard
Lönnig 2011 |